霜黴目的1科。菌絲體發達,少數可形成吸器。多數種的孢囊梗與菌絲無區別,少數具定形的孢囊梗並可繼續生長。孢子囊形態多樣,從線形至球形;萌發生遊動孢子或芽管;遊動孢子腎形,側生等長的雙鞭毛。有性生殖為霜黴目的一般模式,即藏卵器由側生或圍生的雄器授精後形成厚壁的卵孢子。少數可行孤雌生殖。
腐黴科為霜黴目中最原始的1類,是水黴目進化到霜黴目的過渡類型,其中低等類型接近水黴,高等類型接近霜黴。低等種類生於水中或土中,孢子囊不脫落,,與菌絲分化不明顯,萌發時多形成遊動孢子;高等種類陸生,孢子囊易脫落,萌發時產生遊動孢子或直接生芽管。腐黴科腐生於水體或土壤中,或寄生於植物。寄生類型具不同程度的寄生性,一般低等的為弱寄生,寄生范圍較廣;高等的接近專性寄生,有一定的生理分化,但不如霜黴科專化。
腐黴科有些種的“性”似乎是相對的;在性器官和卵孢子的發育中,可能包括與綿黴類似的性激素機制。多數現代研究者認為本科的減數分裂是在配子囊內發生。
本科的模式屬腐黴屬能夠較好地利用六碳糖,除遊離氮和亞硝酸鹽外,其他的含氮物均可作為它的氮源。它能用硝酸鈉合成15種以上的氨基酸,其中最豐富的是丙氨酸和賴氨酸;腐黴對無機硫和有機硫均能利用,並能合成多種維生素,如核黃素、生物素、葉酸、泛酸和抗壞血酸等;它還能分泌果膠酶、纖維素酶以及轉化甾族化合物等。
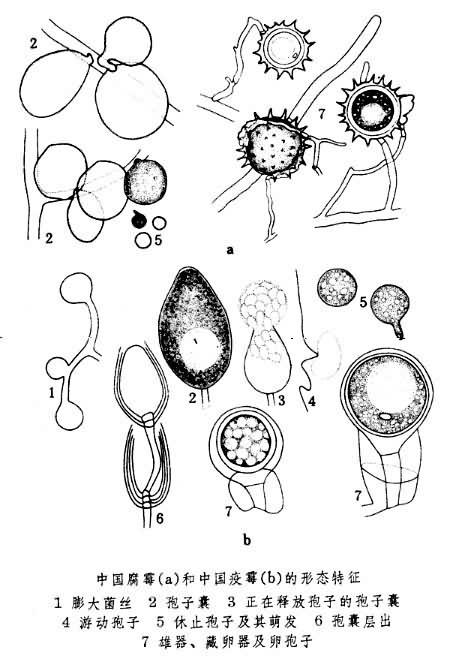
腐黴科雖比水黴科的結構更為復雜,但它與水黴科十分相似,所以早期學者J.施勒特爾(1897)和G.M.史密斯(1938)將它歸入水黴目。後來,F.T.沃爾夫(1947)和F.莫羅等人主張將它提升為腐黴目。G.M.沃特豪斯(1973)在討論霜黴目分類時,將腐黴科分為8屬,常見的重要屬有腐黴屬、疫黴屬、指疫黴屬和霜疫黴屬。腐黴屬包括一些寄生於藻類的水生種和許多寄生於顯花植物的土壤寄藏種,全球約120種,中國有40多種,最著名的是引起幼苗猝倒病的瓜果腐黴和廣泛分佈在中國土壤中的中國腐黴(見圖)。疫黴屬與腐黴屬的主要區別是它的遊動孢子不在孢囊內形成,而是在孢子囊內分化形成。此屬約50種,中國有25種。許多種是經濟植物的重要病原菌,如引起馬鈴薯晚疫病的致病疫黴遍及全球馬鈴薯栽培區,它不僅使馬鈴薯的產量降低,而且孕婦吃瞭病薯以後,可能造成胎兒脊椎發育不全;中國疫黴在中國引起黃瓜疫黴病。指疫黴屬兼具指梗黴和疫黴的特征,有4種,常見的為侵染禾本科植物的大孢指疫黴。霜疫黴屬是中國臺灣、廣東等省荔枝果實上的一單種屬,它的高度分化和有限生長的孢囊梗很象霜黴科;而易於培養、具乳突的孢子囊和圍生的雄器,又很象腐黴科,故G.M.沃特豪斯(1973)將它歸入腐黴科,柯文雄等(1978)主張將它從腐黴科分出,獨立成霜疫黴科。