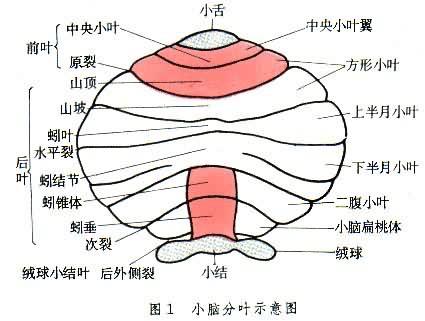
大腦皮層下面、腦幹及腦橋後面的腦組織。小腦可分為前葉、後葉和絨球小結葉(圖1)。
 小腦皮層的結構包括“兩個輸入”(攀緣纖維和苔蘚纖維),5種神經細胞(普氏、籃狀、星狀、高氏和顆粒細胞)和1個輸出(普氏細胞軸突)。除普氏細胞外,其餘4種神經細胞都是中間神經元,它們的輸入和輸出都不越出小腦皮皮層的范圍。顆粒細胞是種族演化上呈現最早,也是中樞神經系統中數量最多的神經細胞。小腦的唯一輸出是投射至小腦深部的小腦核。蚓部投射至頂核,皮層中間部分投射至中間核(人類為齒狀核和栓狀核),外側部位投射至齒核。攀緣纖維和普氏細胞是一對一的聯系,但接觸點則約有300處,是全腦中單個神經細胞和單個輸入纖維間建立的最大數目的突觸聯系。普氏細胞是小腦皮層的樞紐元件。人的小腦皮層的普氏細胞數目約為15×
10
6。一個典型的普氏細胞可與20萬個顆粒細胞的軸突(平行纖維)發生突觸聯系,因此被認為是中樞神經系統中突觸聯系最復雜的神經細胞。普氏細胞樹突的排列,象一片被壓扁的樹冠枝杈伸展在一單個平面中(圖2)。
小腦皮層的結構包括“兩個輸入”(攀緣纖維和苔蘚纖維),5種神經細胞(普氏、籃狀、星狀、高氏和顆粒細胞)和1個輸出(普氏細胞軸突)。除普氏細胞外,其餘4種神經細胞都是中間神經元,它們的輸入和輸出都不越出小腦皮皮層的范圍。顆粒細胞是種族演化上呈現最早,也是中樞神經系統中數量最多的神經細胞。小腦的唯一輸出是投射至小腦深部的小腦核。蚓部投射至頂核,皮層中間部分投射至中間核(人類為齒狀核和栓狀核),外側部位投射至齒核。攀緣纖維和普氏細胞是一對一的聯系,但接觸點則約有300處,是全腦中單個神經細胞和單個輸入纖維間建立的最大數目的突觸聯系。普氏細胞是小腦皮層的樞紐元件。人的小腦皮層的普氏細胞數目約為15×
10
6。一個典型的普氏細胞可與20萬個顆粒細胞的軸突(平行纖維)發生突觸聯系,因此被認為是中樞神經系統中突觸聯系最復雜的神經細胞。普氏細胞樹突的排列,象一片被壓扁的樹冠枝杈伸展在一單個平面中(圖2)。
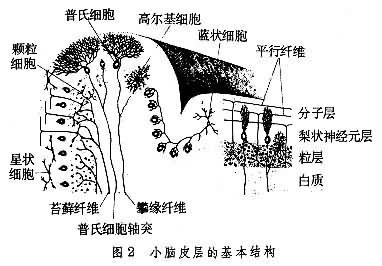 因此平行排列的普氏細胞的樹突,象一片片平行排列的“蜘網”,以捕捉盡可能多的信息。每一個“蜘網”都是一個獨立的系統。由於顆粒細胞的軸突(平行纖維)和壓扁的普氏細胞的“蜘網”成90°角的突觸聯系,又由於籃狀細胞和星狀細胞的軸突既與平行纖維成90°角,又與普氏細胞的軸成90°角,因此小腦皮層的三維晶格結構,被認為是中樞神經系統中罕見的一種“類晶態”結構。
因此平行排列的普氏細胞的樹突,象一片片平行排列的“蜘網”,以捕捉盡可能多的信息。每一個“蜘網”都是一個獨立的系統。由於顆粒細胞的軸突(平行纖維)和壓扁的普氏細胞的“蜘網”成90°角的突觸聯系,又由於籃狀細胞和星狀細胞的軸突既與平行纖維成90°角,又與普氏細胞的軸成90°角,因此小腦皮層的三維晶格結構,被認為是中樞神經系統中罕見的一種“類晶態”結構。
從神經生理來分析,已知小腦皮層的兩個輸入(攀緣纖維和苔蘚纖維)都是興奮性的,小腦皮層的唯一輸出(普氏細胞軸突)則是抑制性的。中間神經元中僅顆粒細胞是興奮性的,而籃狀、星狀和高氏細胞則是抑制性的。在中樞神經系統的任何其他區域,從未發現類似抑制占壓倒優勢的區域。截除全部大腦皮層的哺乳類動物都能自發走動,甚至能避開障礙物;丘腦以上截除的貓和犬也能行走,但躲避障礙物困難;中腦以上截除的貓和犬雖然不能自發走動,但可誘發產生規律的“踏步自動作用”運動。隻要微量(3立方毫米)切除或冷凍(5~0℃)豚鼠小腦蚓部山頂1~2分種,便可使原來的踏步增快至跑步或飛奔。相反,隻要微量(0.3~1毫安)電刺激小腦蚓部山頂幾秒鐘,便可使原來的飛奔降低至跑步或踏步。這些都說明小腦蚓部山頂和中央原始小葉對運動起著抑制性的調控作用。此外,小腦的人原始小葉還有不同的功能定位,例如中央調控後肢、山頂調控前肢、單行調控頸及頭部等。部分切除或可逆冷凍僅能調控,而不能觸發“踏步自動作用”。這種調控作用對自動化程度差的弱“踏步自動作用”特別明顯,而對自動化程度高的強“踏步自動作用”則未必顯著。
小腦主要是調控快或“發射”的運動。研究還發現,小腦深部的小腦核和基底神經節的若幹部位是中樞運動控制的“啟步者”。小腦對運動的調控兼有“開環”和“閉環”兩方面的特性。前者通過存儲在中樞的運動程序或指令進行控制而無需反饋,後者則基於反饋和前饋。例如籃狀細胞對普氏細胞的抑制是一種前饋抑制,而經高氏細胞的抑制則是一種反饋抑制。但是大部分完美的熟練運動和技巧是通過“開環”在無意識中進行控制的。
臨床研究也表示,即使大量切除人的小腦半球的外側部位,也未見明顯的神經學癥狀。但若幹小腦部位的切除或損傷可導致不同表現的運動失調或肌肉群的共濟失調。例如損傷前庭小腦束容易跌倒和引起動暈;損傷脊髓小腦束雖未見癱瘓,但可見牽張反射增強和運動失調。損傷小腦深部小腦核的臨床癥狀最為嚴重,包括有意震顫、解體運動、動幅障礙和運動水平偏差等。例如吃飯時將食物指向鼻孔,走路時抬腿過高和執行較精細運動時震顫特別增強等。
小腦的電生理研究發現,其腦電節律具有明顯的“不活動反應”特性,頻率為150~250赫,比大腦快頻率β節律高5~8倍,但幾乎不因任何感覺輸入的刺激而變化。微電極的研究也表明小腦皮層最主要的普氏細胞的復合峰波也具有罕見的“不活動反應”。
小腦運動控制的化學環節也有明顯的特點。一般認為小腦缺乏多巴胺和P物質,但其3′,5′-環化鳥苷單磷酸(cGMP)和 α-氨基酸氧化酶(α-AAO)的含量特別高,谷氨酸鹽和γ-氨基丁酸 (GABA)的含量也很豐富。α-AAO是一種黃素酶,在哺乳類中樞神經系統中明顯地集中在菱腦部位。雖然該部位僅占全腦重的25%,但 α-AAO的含量超過全腦的90%,並且特別在小腦皮層的顆粒層含量最高。谷氨酸鹽是小腦的興奮性遞質,L-谷氨酸鹽較D-谷氨酸鹽有更強的興奮作用,其含量的增加導致小腦抑制作用的減弱。GABA是小腦皮層的抑制性遞質。普氏細胞至深部小腦核的軸突末梢中含有高濃度的GABA和谷氨酸鹽脫羧酶(GAD),其含量的增高是小腦抑制占壓倒優勢的物質基礎。cGMP在小腦蚓部的含量最高,被認為是小腦皮層動態活動的重要指示器。小腦cGMP的水平由小腦皮層的雙重輸入(苔蘚纖維和攀緣纖維)的活動調節。