在多種無脊椎動物生活環中有不同形態和不同生殖方式個體輪番出現的現象。有的交替過程表現為不同個體一代換一代;但也有的是一種個體在延綿若幹代之後才被另種個體所代替,這種現象稱異態交替。交替的世代中有一代為無性個體而另一代為有性個體,稱無性世代與有性世代交替;一代為單性世代另一代為兩性世代則稱異性世代交替。
原生動物的世代交替 實為有性生殖世代和無性生殖世代的交替。多細胞胞動物交替的各代個體的形成都要經過細胞分裂、分化和生長等階段,可是原生動物的各個世代隻是細胞分裂的直接產物。這是世代交替的低級形式。
在原生動物的世代交替中,交替的有性個體和無性個體並不一定都與單倍體世代和二倍體世代的交替有相應關系;而且有性世代或無性世代可以同屬於單倍體世代或同屬於二倍體世代。據此,原生動物的世代交替可分為下列3型(圖1)。
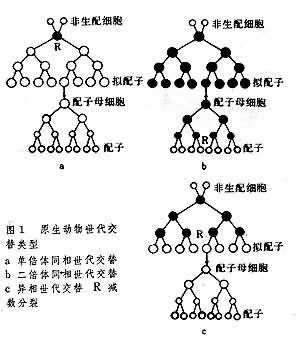
單倍體同相世代交替 有性世代和無性世代的個體都是單倍體,隻有非生配細胞為二倍體。此型見於衣藻和某些胞子蟲類動物。
二倍體同相世代交替 除配子時期為單倍體外,所有無性世代和有性世代的個體都屬二倍體。見於某些太陽蟲和纖毛蟲類動物。
異相世代交替 無性世代為二倍體,有性世代為單倍體,與蕨類植物中胞子體為二倍體配子體為單倍體相一致。有不少有孔蟲類動物屬此型。
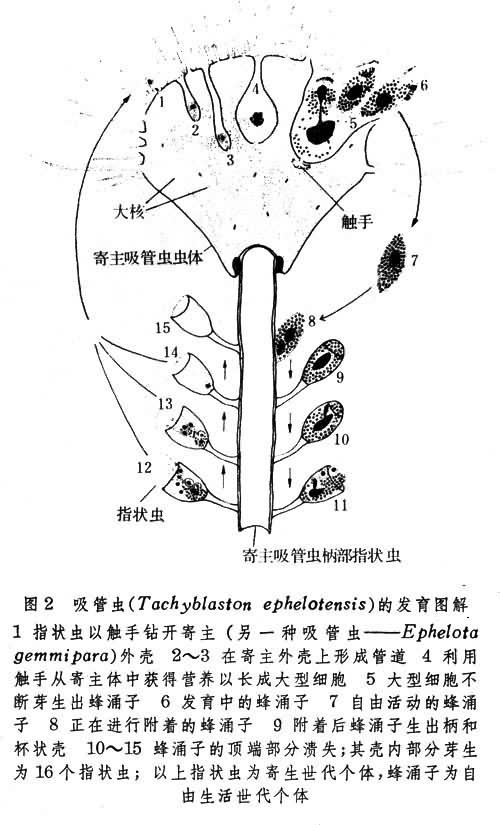
在原生動物中,僅有極個別種類──吸管蟲(Tachy-blaston ephelotensis) 進行有無性世代與另一無性世代的交替。這種動物的指狀蟲寄生於另一種吸管蟲中(Ephelotagemmipara)的胞體中。通過無性生殖(圖2),此蟲體可以不斷地生出蜂湧子,然後再由蜂湧子尋得其他寄主或在本寄主內以出芽法生出下一代指狀蟲。指狀蟲是營寄生生活的無性世代個體,而蜂湧子則是營自由生活的無性世代個體。
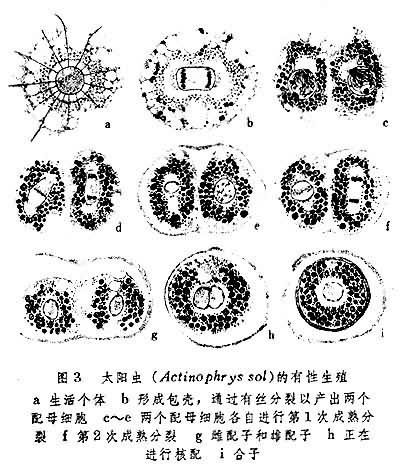
此外,在原生動物中還有兼性世代交替和專性世代交替之分。如太陽蟲(Actinophrys sol) 的世代交替即屬前者。在正常情況下,這種動物進行無性生殖,隻是由於環境的改變,才出現自配形式的有性世代。所謂自配(圖3),就是由個體先形成外面的包殼,然後在殼內通過配母細胞階段以生成雌雄兩個配子。這兩個配子的結合實際上體現為兩個細胞核的結合,因之稱為核配。
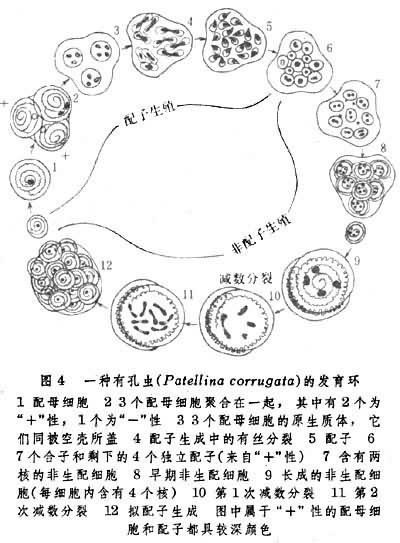
專性世代交替的要點是:由前一世代向後一世代的過渡受體內條件所控制,嚴格按照規定進行,由前一代細胞長大後所產生的子細胞必定改取另一種生殖方法生出再下代,如一種有孔蟲(Patellina corrugta)世代交替(圖4)。這種動物的有性世代是從配母細胞及其結合開始,之後,通過有絲分裂生成一定數量配子。不同的性表現的配子分別結合為定數合子。至此,有性世代結束而無性孔世代──非配子生殖世代開始。這時由各合子發育成的非生配細胞即成體有孔蟲。通過減數分裂和有絲分裂,每個成體可生成一定數量非配子細胞。此種細胞再經長大即成為配母細胞,於是新的一輪世代交替開始。
以上有關兼性世代交替和專性世代交替的概念也適用於多細胞無脊椎動物。
無性世代與有性世代的交替 如腔腸動物的水螅體世代和水母體世代的交替。營固著生活的水螅體為無性世代,營自由生活的水母體為有性世代。在腔腸動物所屬各綱目中,這兩個世代的表現並不一致。有些水螅蟲的水螅體世代占優勢,而另一些水螅蟲和真水母類動物卻在水母體世代得到發展,其水螅體世代表現退化。
水螅體和水母體這兩個世代與動物變態中的幼蟲和成體一樣,雖然有相同的基因型,但表型不同。所以能一時期生成水螅體,另一時期又生成水母體。在此情形下,盡管動物變態中的幼蟲和成體屬同一世代,而世代交替中的水螅體和水母體分屬不同世代,但兩者卻具有相同的遺傳學本質。這一點對追尋世代交替的起源頗有啟發。
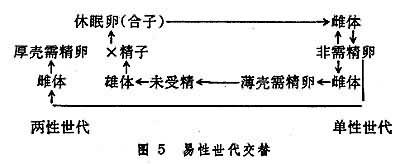
易性世代交替 如有些單巢類輪蟲的單性世代是由休眠卵(或冬卵)發育成的雌性個性。這種個體在春夏間隻能產生不需要受精即能發育的非需精卵並以此重復若幹世代。之後,在季節環境變換影響下,非需精卵可以產生出二種雌體。一種雌體產生的厚殼需精卵,經受精後形成休眠卵,另一種雌體產生的薄殼需精卵不經受精而發育為雄體。於是此雄體與上述產厚殼需精卵的雌體共同形成兩性世代(圖5)。
易性世代交替還常見於昆蟲,如癭蠅類動物(Mias-tor,Obligarces,Micromathus) 的單性世代個體不僅呈幼蟲狀,而且以幼體生殖傳代,以至此種個體雖屬實際上的雌體卻常被稱為幼體生幼蟲。當此單性世代向兩性世代過渡時,首先是蛹化幼蟲的出現。之後再由這種幼蟲分別生出能產雌體的幼蟲和能產雄體的幼蟲,以此進入兩性世代。
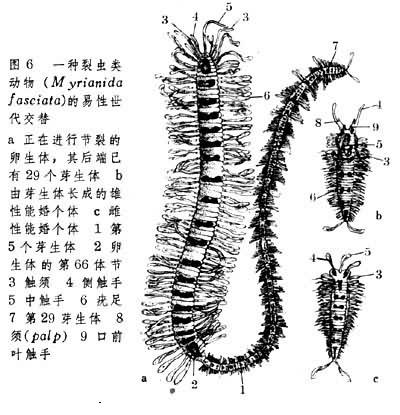
一些多毛類動物(Polychaetes)和棒線蟲(Rhabdia-soidea) 的易性世代交替另有特點。如一種雌雄同體裂蟲(Myrianida fasciata),經交配後產生的受精卵發育成卵生體,此體能通過節體生殖芽生出近30個幼小個體(圖6),是稱芽生體。這些通過無性生殖出來芽生體在脫離親體(卵生體)後分別發育為雌性個體和雄性個體。這時它們又統稱為能婚個體。由能婚個體再產生出卵生體後代。在龍介蟲(Salmacina incrustans)易性世代交替過程中,可註意的是:當進行節體生殖時有的卵生體已具雌雄性腺,有的尚未生成。這樣一來,在前種情形下所形成的交替為雌雄同體兩性世代和雌雄異性兩性世代的交替;而在後種情形下所形成的交替相當於無性世代與有性世代的交替。
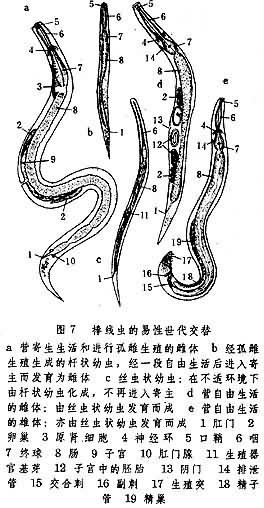
棒線蟲的易性世代交替情況又有不同,其寄生期個體可以是雄性先熟的雌雄同體個體,也可以是進行孤雌生殖的雌體。從這一基礎出發,在環境條件影響下,其世代交替過程會出現兩種情形。一是通過孤雌生殖,由親體產出的卵子孵化為自由生活的幼蟲,然後由幼蟲進入寄主以發育為新一代寄生個體。這種與親代相同的新個體被稱為棒線蟲的殖同世代;由親代到此世代生成的全過程稱為殖同生活環。二是通過孤雌生殖產出的幼蟲並不直接進入寄主而是分別生成雌性個體和雄性個體。由此兩性個體產生的新一代個體再恢復寄生生活。由於以上兩性個體被稱為棒線蟲的殖異世代,所以由親體產生此世代以及由此世代產生下一代寄生個體的全過程被稱為殖異生活環。棒線蟲殖同世代與殖異世代的交替實際上也是兩個生活環的交替(圖7)。
單倍體世代與二倍體世代的交替 這是以染色體數在生活環中的變換為基礎,在放寬世代交替含意下所提出的一個概念〔見世代交替(植物)〕。任何真核生物都有單倍體和二倍體這兩個時期或世代,其交替過程是:由二倍體細胞通過減數分裂以生成配子形式的單倍體,然後經過配子的結合再恢復為雙倍染色體數的二倍體。對真菌來說,有絲分裂限在單倍體世代進行,但對高等植物來說,以上兩個世代都可進行有絲分裂,隻是其規模各有不同,從而形成不同發育程度的配子體和胞子體。其中配子體是單倍體世代,胞子體是二倍體世代。動物界和高等植物都是二倍體世代占絕對優勢,真菌類則是單倍體占絕對優勢。
從進化觀點來看,二倍體較之單倍體更具優越性。原因是經異型接合形成的二倍體一方面可以收藏或儲存許多一時不起作用並在單倍體中就會丟失的基因,從而使基因組合的潛在變異能力得以提高,有利於通過遺傳機制產出更具適應能力的新生物種;另一方面由於隱性基因表現機會少、傳播緩慢,有緩解自然選擇壓力的作用,所以生物體就可避免來不及對突然選擇作出快速應變。反之,如果沒有隱性基因的這一作用,生物體的變異將更加劇烈。
環境條件與世代交替的關系 太陽蟲在缺乏食物時才會出現有性世代。海月水母有性世代(以碟狀體為開始)的生成與海水中化學元素(碘)濃度有密切關系(見生活環(動物))。枝角類動物兩性世代的出現取決於水體溫度變化和幹涸程度。不少輪形動物(Asplanchna,Brachionus,Testudinella)在培養液或食物更換時,或在pH或碳酸鹽濃度有所升降時,都會生出能產需精卵的雌體。以此導致兩性世代的到來。
癭蠅的幼體生幼蟲(即單性世代的雌體)向雌雄兩性世代的過渡實際上也要受到許多環境條件的影響。當營養條件良好時,其單性世代可延綿不斷。反之,如有饑餓、強光照射、幹燥或擁擠發生,作為向兩性世代過渡期的蛹化幼蟲即在下代出現。不過這種幼蟲有很明顯的不穩定性,即仍處於發育方向尚未完全決定狀態。譬如幼蟲的出現是由饑餓造成,那麼,在重獲豐富食物之後,就會發生還童現象(見生活環(動物))。這時的蛹化幼蟲無論在形態上或在生理上又都恢復到幼體生幼蟲水平。
再如寄生原生動物中,有些胞子蟲的生活環中有3個世代即:裂殖生殖、配子生殖和胞子生殖。在配子生殖未到之前,裂殖生殖可以重復若幹代,其數目的多少全由寄主受染器官所供營養和空間條件而定(圖8)。至於配子生殖向胞子生殖的過渡,一切要受內部條件控制,必須按一定的發育順序和規律進行。所以,由裂殖生殖到配子生殖的交替具有兼性世代交替性質,而由配子生殖到胞子生殖的交替卻具專性世代交替性質。
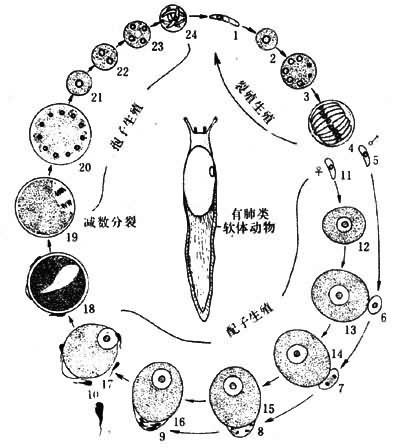 圖8 一種裂球蟲(Klossia)的發育環
圖8 一種裂球蟲(Klossia)的發育環