含磷酸的複合脂質。包括磷酸甘油酯(又稱甘油磷酸酯)和鞘磷脂兩類。生物體的重要組分,如動物的腦、肝、紅細胞和卵黃等以及植物的種子含量較多,磷脂是細胞膜和各種細胞器(線粒體、內質網、細胞核、高爾基器、葉綠體等)膜的重要組分,幾乎細胞所含有的全部磷脂都集中在生物膜中。生物膜的許多特性,如作為膜內外物質的通透性屏障,膜內外物質的交換,資訊傳遞,神經脈衝的傳導等都與磷脂和其他膜脂有關。磷酸甘油酯的主鏈是甘油,甘油的第三個羥基被磷酸酯化,另外兩個羥基被脂肪酸酯化,磷酸酸基團又與各種結構不同的小分子化合物相連接。兩個長碳氫鏈(脂肪酸鏈)具有非極性特性,甘油分子的第三個羥基與磷酸形成的酯鍵是有極性的;所以這類化合物是親水脂兩性分子。常見的磷酸甘油酯有磷脂酰膽堿(卵磷脂)、磷脂酰乙醇胺(腦磷脂)等。鞘磷脂的主鏈是鞘氨醇(含氨基的長鏈醇類化合物),脂肪酸以酰胺鍵連接在它的氨基上,磷酸以酯鍵連接在它的1-羥基上。鞘磷脂也是親水脂兩性分子,是高等動物神經組織中含量最豐富的鞘脂類(鞘氨醇是鞘磷脂的主要成分,故亦屬於鞘脂類)。磷脂能在生物體內合成並快速地周轉。
結構及命名 磷酸甘油酯 甘油分子的中央碳原子是不對稱的。天然的磷酸甘油酯都具有相同的立體化學構型,屬於L系(見圖)。
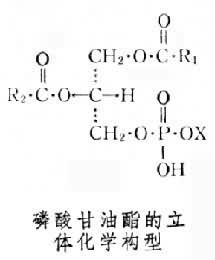 根據IUPAC-IUB國際委員會制定的脂質命名原則,磷酸甘油酯中:如X為膽堿,則應命名為:1,2-二酰基-sn-甘油-3-磷酰膽堿,亦稱L-3-磷脂膽堿,俗名卵磷脂。圖上構型中
R
1,
R
2代表脂肪酸鏈,X為連接在磷酸上的小分子化合物;名稱中sn為立體化學專一編號。
根據IUPAC-IUB國際委員會制定的脂質命名原則,磷酸甘油酯中:如X為膽堿,則應命名為:1,2-二酰基-sn-甘油-3-磷酰膽堿,亦稱L-3-磷脂膽堿,俗名卵磷脂。圖上構型中
R
1,
R
2代表脂肪酸鏈,X為連接在磷酸上的小分子化合物;名稱中sn為立體化學專一編號。
磷酸甘油酯分子內部既含有強極性基團同時也含有強非極性基團。兩個脂肪酸鏈形成非極性尾,而含磷酸的一端是極性頭部。各種磷酸甘油酯的差別主要在於其極性頭的大小、形狀和電荷的差異。L-磷脂酸是最簡單的磷酸甘油酯,磷酸基團上不連接任何小分子化合物。它是各種磷酸甘油酯的母體化合物,廣泛地存在於細胞內,但僅有痕量,因為周轉率很快,是合成各種磷脂和脂肪的關鍵中間產物。
每一種磷酸甘油酯都不是單純的化合物,如磷脂酰膽堿分子內脂肪酸組成就是多種多樣的。絕大多數磷酸甘油酯C-1位上以飽和脂肪酸為主,而C-2位上不飽和脂肪酸居多。
磷酸甘油酯分子中的碳氫鏈並不是無例外地以酯鍵連接在甘油的羥基上。縮醛磷脂的甘油分子中第一個碳原子由順式烯醚鍵連接碳氫鏈,第二個碳原子以酯鍵連接長鏈脂肪酸。極性頭通常是乙醇胺。另外還有一種醚磷脂是縮醛磷脂的還原產物,甘油分子的C-1以醚的結構連接碳氫鏈,這種化合物比較罕見。
鞘磷脂 鞘磷脂與磷酸甘油酯的差別在於脂肪酸殘基是連接在鞘氨醇的氨基上,“X”基團是通過磷酸連接到鞘氨醇的C-1羥基。“X”通常為膽堿或乙醇胺。鞘磷脂分子內的鞘氨醇碳鏈和脂肪酸碳鏈形成非極性尾,含“X”的磷酸端為極性頭,也是親水脂兩性分子。神經組織鞘磷脂內的脂肪酸限於硬脂酸、廿四烷酸和神經酸。脾臟和肺臟鞘磷脂內的脂肪酸主要是棕櫚酸和廿四烷酸。長鏈鞘氨醇有兩類:鞘氨醇型和4-羥基雙氫鞘氨醇型(亦稱植物鞘氨醇型)。各種不同的鞘氨醇的差別在於
表1 生物體鞘脂內的鞘氨醇類型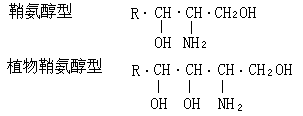 碳鏈長短(
C
14~
C
24);雙鍵數目與構型;碳鏈分支(異-和反異-)生物體含有各種不同的長鏈鞘氨醇,在高等動物中,依進化趨勢其碳鏈加長,不飽和度增加;植物和真菌的長鏈鞘氨醇含有三個羥基;海洋無脊椎動物以雙不飽和化合物為主(表1)。
表1 生物體鞘脂內的鞘氨醇類型
碳鏈長短(
C
14~
C
24);雙鍵數目與構型;碳鏈分支(異-和反異-)生物體含有各種不同的長鏈鞘氨醇,在高等動物中,依進化趨勢其碳鏈加長,不飽和度增加;植物和真菌的長鏈鞘氨醇含有三個羥基;海洋無脊椎動物以雙不飽和化合物為主(表1)。
表1 生物體鞘脂內的鞘氨醇類型

磷酸甘油酯的性質及類別 純的磷酸甘油酯是白色蠟狀固體,暴露在空氣中變為棕紅或棕黑色,這是因為其中的不飽和脂肪酸被氧化形成過氧化物的聚合物之故。磷酸甘油酯能溶於含少量水的非極性溶劑;與水相混合自動形成微團或片狀雙層結構,親水的極性頭面向水相,而疏水尾則互相聚集於微團內側。這種脂質(磷脂和少量糖脂及膽固醇)雙層是生物膜的基本結構。各種磷酸甘油酯所含極性頭的大小,形狀和電荷都不一樣。磷脂酰膽堿,磷脂酰乙醇胺是既帶有正電荷又帶有負電荷的兩性離子;磷脂酰絲氨酸分子在pH=7時帶有兩個負電荷和一個正電荷。這三者結構相似,親水性強。具有強親水極性頭和疏水尾結構的化合物都有優良的去垢作用。表2列出磷酸甘油酯的極性頭結構類別,名稱及在生物體的分佈。縮醛磷脂在肌肉和神經細胞膜中含量豐富。醚磷脂比較罕見,存在於某些種動物的紅細胞及蛞蝓體內。腦磷脂是一個舊的俗稱,指由磷脂酰乙醇胺和磷脂酰絲氨酸以及少量其他脂質組成的混合物。
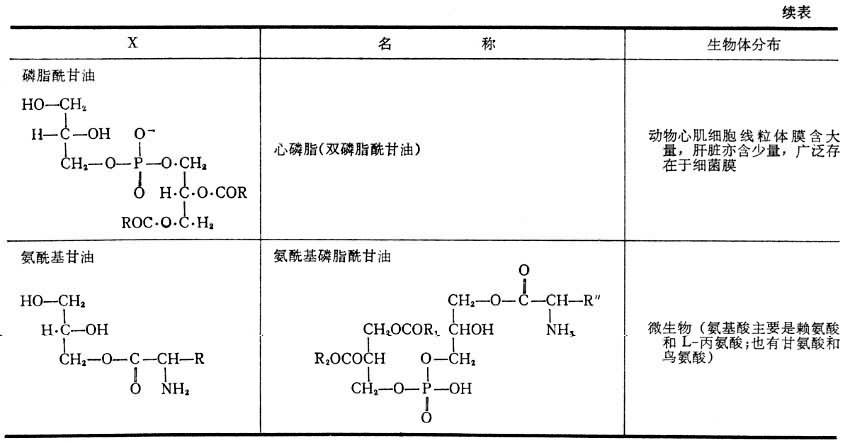
表2 磷酸甘油酯的極性頭結構類別、名稱及生物體內的分佈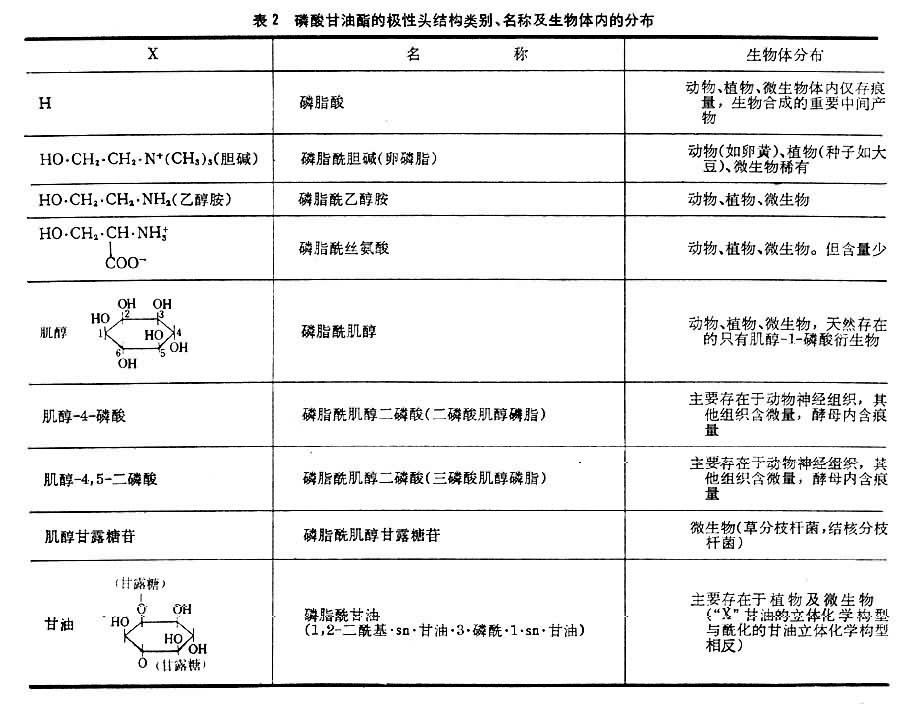 表2 磷酸甘油酯的極性頭結構類別、名稱及生物體內的分佈 續表
表2 磷酸甘油酯的極性頭結構類別、名稱及生物體內的分佈 續表
磷脂的生物合成 磷脂酸是合成磷脂的前體,同時也是合成三酰甘油的前體。有幾條不同的途徑生成磷脂酸。
磷脂酸經兩條途徑合成不同的磷脂,兩者都利用胞苷三磷酸輔因子。其一是磷脂酸經磷脂酸磷酸酯酶水解產生二酰甘油,後者與胞苷二磷酸膽堿或胞苷二磷酸乙醇胺作用產生磷脂酰膽堿或磷脂酰乙醇胺。在這裡,胞苷二磷酸是“堿基”(X)的載體。全程合成始於膽堿或乙醇胺的磷酸化,活化的膽堿或乙醇胺與胞苷三磷酸作用形成CDP-膽堿或CDP-乙醇胺。
①甘油磷酸途徑

②脂酰基磷酸二羥丙酮途徑

③甘油二酯激酶途徑
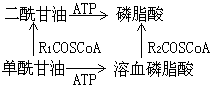
另一途徑是磷脂酸與胞苷三磷酸反應生成“活化的磷脂酸”即胞苷二磷酸二酰甘油,後者能與絲氨酸或肌醇作用分別合成磷脂酰絲氨酸(細菌合成途徑)或磷脂酰肌醇。磷脂酰肌醇經磷酸化反應能產生二磷酸肌醇磷脂和三磷酸肌醇磷脂(動物腦)。胞苷二磷酸二酰甘油還能與sn-甘油-3-磷酸反應產生3-sn-磷脂酰-1'-sn-甘油-3'-磷酸,再經水解切除磷酸基團從而合成磷脂酰甘油。後者再與CDP-二酰甘油作用就合成瞭心磷脂(動物體)。細菌利用2分子磷脂酰甘油縮合合成心磷脂。
動物體合成磷脂酰絲氨酸是通過酶促交換極性頭的反應。動物體也能由磷脂酰乙醇胺的直接甲基化合成磷脂酰膽堿。磷脂酰絲氨酸經脫羧又能轉變成磷脂酰乙醇胺。極性頭的交換反應和磷脂之間的相互轉變在改變膜磷脂的極性頭分子結構上起重要作用。
鞘氨醇的氨基被長鏈脂肪酸酰化產生的N-脂酰鞘氨醇(神經酰胺)與CDP膽堿作用合成鞘磷脂。
磷脂的酶促降解及生理功能 磷酸甘油酯在生物體內經磷脂酶水解成為甘油、脂肪酸、磷酸和各種小分子化合物(X)。磷脂酶A1,A2,B,C,D分別作用於磷酸甘油酯分子內的不同酯鍵。
磷酸甘油酯的降解代謝通常不一定進行到底,部分降解的中間產物還能被細胞重新利用。對細胞來說,重新利用中間產物一方面節省能量,不必一切都從頭合成;另一方面也能按照機體的需要改制磷脂分子。
催化磷脂降解的各種酶在磷脂周轉和改造中起重要作用。如磷脂酶 A(包括A1和A2)能催化產生小量溶血磷脂,它們和酰基轉移酶協同工作,對機體脂質內脂肪酸組成的調節改組起重要作用,是機體適應細胞需要“定做”具有特定脂肪酸成分的脂質的主要方式之一。磷脂酶A2能從膜磷脂釋放花生四烯酸作為“在位”合成前列腺素的前體。磷脂酶A催化產生溶血磷脂富集的膜區,從而改變膜的通透性屏障。磷脂酶和酰基轉移酶還能修復和更新那些由於自身氧化或其他損傷而造成的劣質磷脂分子,維持膜磷脂的完整。