導致生殖母細胞中染色體數目減半的分裂過程(見彩圖)。它是1883年由 E.van貝內登在動物和1888年由E.A.施特拉斯佈格在植物最初發現的。在所有進行有性生殖的生物的生活史中,細胞除進行數次有絲分裂外,還要進行一次減數分裂,使其染色體數目由二倍體(2n)變為單倍體(n)。減數分裂發生在配子形成前的某一時期,所以雌雄配子的核都是單倍的。受精後形成的合合子又成為二倍的。由於減數分裂,使每種生物代代都能夠保持二倍體的染色體數目。在減數分裂過程中非同源染色體重新組合,同源染色體間發生部分交換,結果使配子的遺傳基礎多樣化,使後代對環境條件的變化有更大的適應性。動物的減數分裂發生在形成配子之前。在雄性個體的精細管中,每個精母細胞通過減數分裂形成四個精細胞,精細胞分化成為精子。在雌性個體的卵巢中,卵母細胞通過減數分裂形成卵細胞。被子植物、裸子植物、蕨類和苔蘚植物的生活史都由孢子體和配子體世代組成,減數分裂發生在孢子體世代的末尾。在被子植物的雄蕊花藥中,小孢子母細胞通過減數分裂形成四個小孢子。小孢子經有絲分裂發育成雄配子體。雌蕊子房中的大孢子母細胞通過減數分裂形成大孢子。大孢子經有絲分裂發育成雌配子體。藻類和菌類進行減數分裂的時期依物種的不同而有多種情況。極端特殊情況見於水綿和衣藻,它們的配子結合(受精)形成合子,合子的第一次分裂就是減數分裂(見有性生殖)。
 細線期
細線期
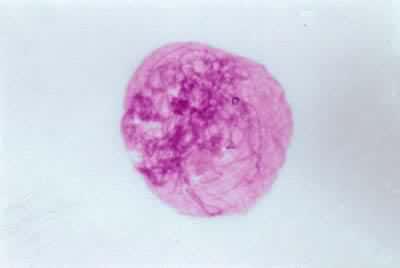 偶線期
偶線期
 粗線期
粗線期
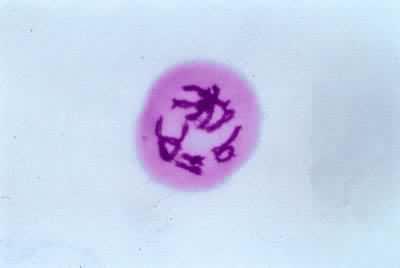 雙線期
雙線期
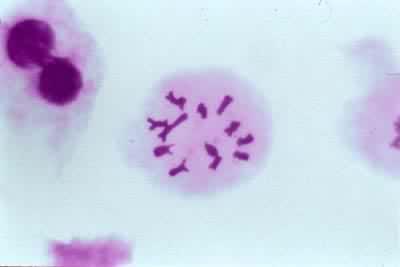 終變期
終變期
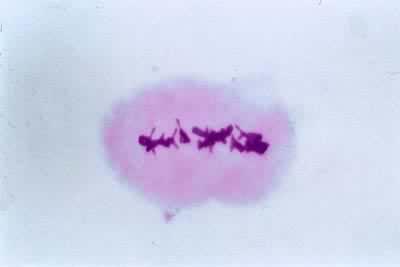 中期Ⅰ
中期Ⅰ
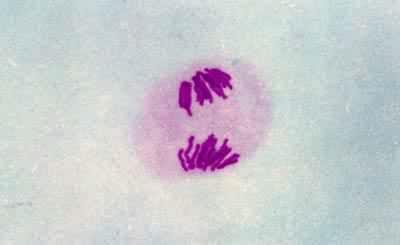 後期Ⅰ
後期Ⅰ
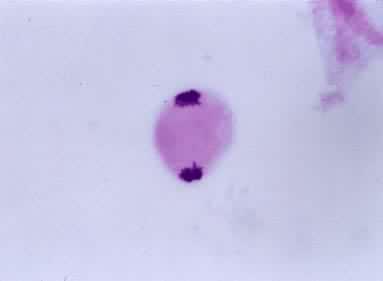 末期Ⅰ
末期Ⅰ
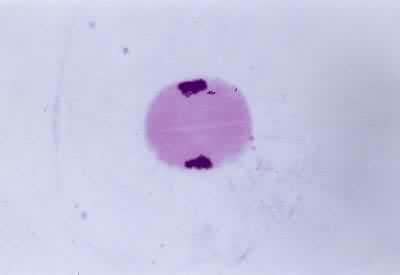 間期
間期
 前期Ⅱ
前期Ⅱ
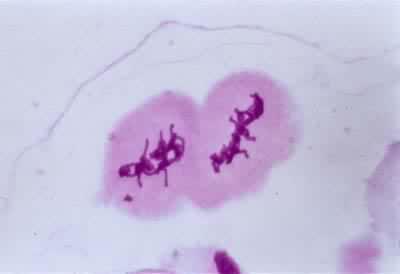 中期Ⅱ
中期Ⅱ
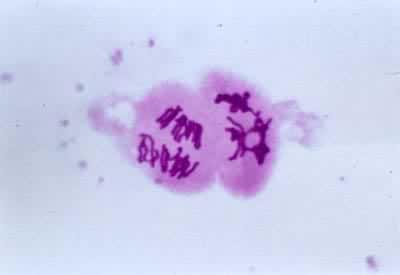 後期Ⅱ
後期Ⅱ
 末期Ⅱ
末期Ⅱ
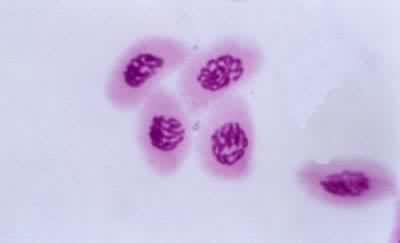 四分子
四分子
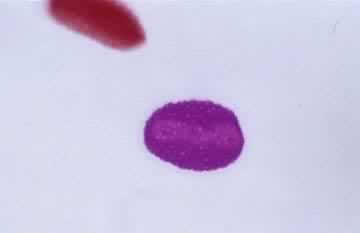 花粉粒
花粉粒
過程 減數分裂是由相繼的兩次分裂組成的,分別稱為減數分裂Ⅰ和減數分裂Ⅱ。在這兩次分裂之間一般有一很短的間期,不進行DNA合成,從而也不發生染色體復制。由於細胞核分裂兩次,而染色體隻復制一次,所以經過減數分裂染色體數目減半。
減數分裂Ⅰ 前期比較復雜,減數分裂的許多特殊過程都發生在這一時期。因是第一次分裂的前期通常稱為前期Ⅰ。它又細分為:①細線期,染色質已集縮成細長的線狀結構,每條染色體通過附著板與核膜相連,局部可見念珠狀小節稱為染色粒。一般認為染色粒是染色線緊密地螺旋折疊的結果。此期核的體積增大,核仁也較大。②合線期亦稱偶線期,是同源染色體配對的時期。這種配對稱為聯會。每對中的兩條同源染色體分別來自雌性和雄性生殖細胞,它們在形態和遺傳結構上是相似的。同源染色體的配對一般是從靠近核膜的一端開始,有時在染色體全長的若幹點上也同時進行聯會。配對是靠兩條同源染色體間沿長軸形成的聯會線復合體實現的。配對後的每對同源染色體稱二價體。由於聯會,細胞中的染色體由2n條單價體成為n條二價體,雖然DNA含量未變,但數目看起來減少瞭一半。③粗線期,染色體明顯縮短變粗。因此,在一些染色體數不多的生物,此時可以看出細胞中有幾個二價體。聯會的兩條同源染色體結合緊密,隻在局部位置上有時可分辨出是兩條染色體。在有些植物,例如玉米,此期二價體的著絲粒、染色粒、異染色質區和核仁組織區都可以看清,因此利用這些特征和染色體長度可做核型分析。在粗線期每條染色體實際已由兩條染色單體組成,來自同一染色體的稱為姐妹染色單體,對它們來說來自另一條染色體的單體則是非姐妹染色單體。在粗線期同源染色體的非姐妹染色單體間發生局部交換。因為聯會非常緊密,這種交換過程不能直接看到。但可根據下一時期(雙線期)看到的交叉現象,以及同源染色體間基因重新組合的遺傳學事實,判斷發生瞭這種過程。粗線期核仁仍然很大,含有很多RNA。④雙線期,聯會的兩條同源染色體開始分離,但在許多稱作交叉的點上它們還連在一起。此期可以看清,聯會的兩條染色體都分別由兩條染色單體組成。故每個二價體由四條染色單體構成。交叉發生在兩條非姊妹染色單體之間。一般認為,交叉是發生瞭交換的結果。雙線期的染色體進一步縮短,此時聯會絲復合體已消失。在植物一般雙線期的時間比細線期、合線期和粗線期都短得多。但人和動物的卵母細胞常長期停留在減數分裂的雙線期。例如人的卵母細胞在五個月的胎兒中已達雙線期,而一直到排卵時都停在此期。⑤終變期亦稱濃縮期,二價體顯著收縮變粗,並向核的周邊移動,在核內較均勻地分散開。核仁此時開始消失,但有的植物,如玉米、水稻,在終變期的早期核仁仍然很大,並染色很深。交叉數目減少,終變期末有些二價體的同源染色體隻在末端連在一起。
中期Ⅰ核膜解體後二價體分散在細胞質中。其中的一條染色體通過牽引絲與一極相連,而另一條染色體則通過牽引絲與另一極相連。最後二價體排列於赤道區,形成赤道板。
後期Ⅰ每個二價體的兩條同源染色體分開,移向兩極。n個二價體成為n條單價染色體,此時DNA含量減半。二價體中那條染色體移向那一極是隨機的。
末期Ⅰ染色體各自到達兩極後逐漸解螺旋化,變成細線狀。核膜重建,核仁重新形成,同時進行細胞質分裂。許多植物在減數分裂Ⅰ隻發生核的分裂,而細胞質分裂在減數分裂Ⅱ的末期進行,使四個核同時分開。
間期 在減數分裂Ⅰ和減數分裂Ⅱ之間的間期很短,不進行染色體復制。這時每條染色體已是由兩條染色單體構成瞭。在有些生物甚至沒有這個間期,而由末期Ⅰ直接轉為前期Ⅱ。
減數分裂Ⅱ 這次分裂基本上與有絲分裂相同。前期Ⅱ時間較短。中期Ⅱ染色體排列於赤道面,兩條染色單體的著絲點分別向著兩極,形成赤道板。後期Ⅱ時兩條染色單體分開,移向兩極。到達兩極的子染色體為n數,並且每條子染色體隻由一條染色單體構成。末期Ⅱ時兩極的子染色體解螺旋化。形成核膜,出現核仁,經過細胞質分裂,完成減數分裂過程。新產生的每個細胞都變成瞭單倍體。
同源染色體的聯會 減數分裂機制的核心是同源染色體的聯會和交換。用電子顯微鏡觀察發現,在配對的同源染色體之間沿縱軸形成聯會線復合體(SC)。靠它把每對同源染色體穩定地聯在一起。SC的細微結構在各種動植物中大體相同。兩側是約400埃寬電子密度很高的側部組分,很暗。兩側之間為約1000埃的低電子密度的中間區,在電子顯微鏡下較為明亮。在中間區的當中為中央組分,約300埃是比較暗的區域。SC的這些部分的厚度依生物種類有些差異。側部組分的外側纖維和染色體基本纖維之間連接十分緊密,以至難以區分。在側部組分和中央組分之間有細纖維相連結,稱L-C纖維。L-C纖維直徑為70~100埃左右。有人推測,L-C纖維在中間區相遇,形成中央組分。此過程稱為鎖合。側部組分主要由蛋白質構成,其中也有少量DNA。L-C纖維和中央組分也主要由蛋白質組成。關於其中是否有DNA尚無確切的證據。關於SC形成的詳細過程還不清楚。有實驗說明,細胞中的DNA大約有0.3%是在合線期合成的。這種DNA稱“合線期DNA”。它並不分佈在染色體的某一位置上,而是散在於整個染色體上。如果在細線期或合線期加入DNA合成抑制劑。抑制合線期DNA合成,則同源染色體不能配對,SC不能形成。如果在合線期阻抑蛋白質的合成,也會使同源染色體的配對過程遭到阻礙,停止SC形成。實驗表明,細胞中存在著許多基因支配聯會過程,有的促進、有的抑制聯會。
交換的機制 曾經提出過各種模型來解釋交換過程的機制,但至今沒有一個被公認。從分子水平理解交換的機理必然包括DNA的斷裂和再結合過程。DNA的切斷必須有核酸酶參加,在百合中發現有一種脫氧核糖核酸酶(最適pH為5.2)在合線期活性開始上升,粗線期達最高峰,而在粗線期末消失。DNA的斷口再結合要有連接酶參加。有實驗表明,由合線期末到粗線期連接酶的活性上升。在許多材料中(百合、小麥、黑麥、蠑螈、小鼠和人)都看到在粗線期有DNA合成。這時合成的DNA稱為“粗線期DNA”(P-DNA)。它與合線期DNA不同,是由非常短的片斷構成的。如果抑制P-DNA的合成,則染色體就會發生斷裂。因此 P-DNA對保持染色體的完整性是很重要的。推測P-DNA在DNA斷裂的再結合過程中對斷口的修復可能起一定作用。在DNA酶的作用下,DNA可產生很多斷口,但同源的二條染色體在相同的位置上都產生斷口的機率是不高的,隻有這樣的斷口有可能發生交換,其他大部分斷口將被修復。同源染色體間實現局部交換,必須在空間上非常靠近,否則交換過程是不可想像的。因此聯會是實現交換的前提條件。有許多實驗證明,抑制聯會過程,不形成SC,則交換過程不能發生,也看不到交叉現象。例如,在合線期抑制蛋白質的合成或合線期DNA的合成,則聯會復合體不能形成,聯會不能進行,於是也不發生交換和交叉。
參考書目
郝水編:《有絲分裂與減數分裂》,高等教育出版社,北京,1983。
B.Alberts,et al.,Molecular Biology of the Cell,Garland Publishing,Inc.,New York &London,1983.