含糖的脂質化合物,其糖(鏈)與脂中的醇基以共價連接。糖脂除參與質膜構成外,還與細胞表面標誌及細胞間的粘合等生物學功能密切相關。
糖脂廣泛存在於各種生物體中。自然界中的糖脂可按其組分中的醇基種類而分為兩大類:甘油糖脂及鞘糖脂。糖基化的甘油醇脂類稱為甘油糖脂,或稱糖脂醯甘油,存在於動物的神經組織、植物和微生物中,是植物中的主要糖脂,亦是某些細菌,尤其是革蘭氏陽性細菌菌膜的常見組成成分。糖基化的鞘氨醇脂類稱為鞘糖脂或糖鞘脂,最最初發現於神經組織中,以後發現在動、植物的各種細胞膜中普遍存在。其生物學功能遠較甘油糖脂復雜。其類型及其與其他各種類脂在組織、細胞中所占的比例因種屬及器官而異。尤其是腎中糖脂的種屬特異性十分突出。僅含中性糖及氨基糖的鞘糖脂稱為中性糖脂。其中隻含一個單糖基的稱為腦苷脂。半乳糖腦苷脂在腦白質、脊髓及腎中含量豐富,而葡萄糖腦苷脂則存在於非神經組織的細胞膜中。腦苷脂去掉脂肪酰則稱為鞘氨醇糖苷。硫酸化的腦苷脂稱為硫酸腦苷脂或腦硫脂。含唾液酸的鞘糖脂稱為神經節苷脂,因最初在腦組織中發現,誤認為是神經節特有的成分而得名,以後證明廣泛存在於神經組織以外的各種細胞及體液中。此外,甘油磷脂或鞘氨醇磷脂亦均可發生糖基化。
糖脂的生物學作用見表1。
表1 細胞表面鞘糖脂的生物學作用
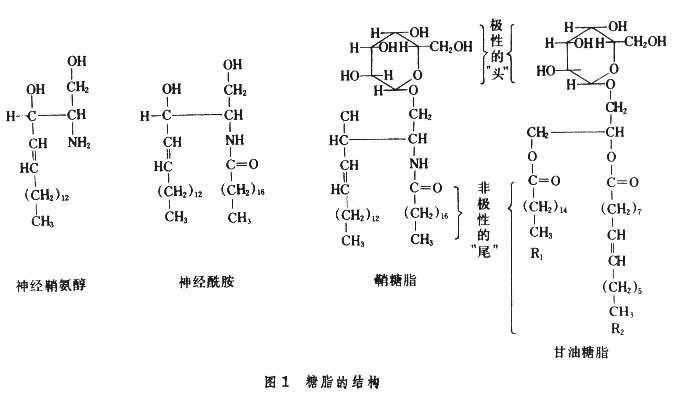
結構、分類及命名 糖脂的組成成分主要是醇、脂肪酸及糖(圖1)。有的還有硫酸或磷酸。甘油糖脂的醇為丙三醇(甘油),與脂肪酸形成酯鍵。鞘糖脂的醇為鞘氨醇(神經鞘氨醇),主要存在於哺乳動物,以其C-2氨基與脂肪酸形成酰胺鍵,產生N-脂酰鞘氨醇,又稱神經酰胺。糖成分一般構成糖鏈(直鏈或有分枝)。糖鏈多以糖苷鍵與醇的羥基連接,分別形成甘油糖脂或鞘糖脂。有的與磷脂酰甘油或磷脂酰鞘氨醇的磷酸基形成酯鍵,還有與肌醇磷脂或肌醇鞘磷脂的肌醇形成糖苷鍵。最簡單的糖脂隻含一個糖基,如半乳糖腦苷脂及葡萄糖腦苷脂。復雜的鞘糖脂可有20個以上糖基組成的糖鏈。鞘糖脂中常見的糖基為D-葡萄糖 (Glc)、D-半乳糖 (Gal)、D-乙酰氨基葡萄糖(GlcNAc)及D-乙酰氨基半乳糖(GalNAC),有的還有L-巖藻糖 (Fuc)、D-甘露糖 (Man)及唾液酸(SA)。不含唾液酸的非硫酸化鞘糖脂稱為中性鞘糖脂,不帶電荷。含唾液酸的鞘糖脂稱為神經節苷脂。神經節苷脂在中樞神經系統含量較高,在灰質中的含量相當於白質中的3倍;在非神經組織中含量雖不高,但功能非常重要,是膜脂不可缺少的成分。神經節苷脂中唾液酸的羧基在生理性pH下解離而使整個分子帶負電荷。
鞘糖脂的某些糖殘基(常為Gal的C-3羥基)可發生硫酸化,成為帶負電荷的硫酸鞘糖脂。它們主要存在於神經組織的髓鞘及白質中。例如硫酸化的半乳糖腦苷脂約占腦白質脂類的15%。
神經節苷脂及硫酸鞘糖脂屬於酸性鞘糖脂。它們根據鞘糖脂糖鏈根部的結構可分為7個不同的系,傳統命名為GM4、GM3、GM2、GM1、GD1a 、GD1b、GT1a 、GT1b及GT1c。
綜上可見,甘油糖脂為糖基化的二脂酰甘油,鞘糖脂為糖基化的單脂酰鞘氨醇。兩者各有兩條非極性的疏水長“尾”(前者由兩個脂肪酰提供,後者由鞘氨醇及脂肪酰各提供一個)。這兩條疏水長“尾”可嵌入於膜脂雙層,由糖基構成的極性“頭”部伸向質膜外,可作為細胞表面標志或抗原決定簇或受體的識別部分。
特性 糖脂屬於兩親(即親水又親脂)性物質。其水溶性差別很大。有的完全不溶於水,如含單糖基的腦苷脂;有的則能以微泡或聚合物形式“溶”於水,如復雜的中性鞘糖脂及神經節苷脂。糖脂分子中由鞘氨醇及脂肪酰烴鏈構成的疏水性長尾通常嵌入於質膜脂雙層的外層,故親水性糖鏈僅暴露於細胞外表面,呈不對稱分佈。
代謝 鞘糖脂的轉換率在中樞神經系統,尤其在新生兒期,十分迅速。
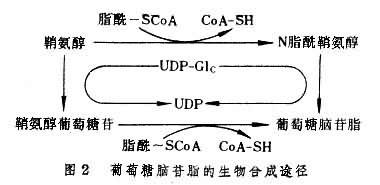
鞘糖脂的合成過程經過多步驟在內質網及戈爾吉氏體中進行。整個合成過程包括鞘氨醇的生成、鞘氨醇脂酰化(神經酰胺的生成)及糖鍵的合成三個階段。鞘氨醇系由脂酰輔酶A與絲氨酸縮合並脫羧,再經還原及黃酶催化的脫氫而生成。鞘氨醇可通過與脂酰輔酶 A或遊離脂肪酸作用而進行脂酰化生成脂酰鞘氨醇(神經酰胺)。脂酰輔酶A參加的反應由脂酰轉移酶催化;遊離脂肪酸參加的反應由一種水解酶催化(即神經酰胺分解的逆反應);一般以前者為主(圖2)。糖基化的第一個產物為葡萄糖腦苷脂或半乳糖腦苷脂。接受第一個糖基的可為脂酰化的或未脂酰化的鞘氨醇。前者直接生成腦苷脂;後者先生成鞘氨醇糖苷,然後再脂酰化生成腦苷脂。單糖基腦苷脂可進一步逐個增加糖基,形成含糖鏈的鞘糖脂。每一步糖基化反應都在一定的糖基轉移酶的催化下進行。這類酶的特異性很強,即對其兩個作用物,單糖供體及糖基接受體,具有雙重特異性。這些酶都是膜結合的或嵌入膜中的,並可能以復合酶體的形式存在。每一種多酶復合體合成一種糖脂。前體一旦結合到復合酶體便按預定順序進行連續糖基化,直至生成特定的糖鏈。糖基化的單糖供體為糖核苷酸。糖脂的硫酸化常發生在Gal基第3位碳原子上,由3'-磷酸腺苷5'-磷酰硫酸(PAPS)提供硫酸,形成硫酸酯鍵。
鞘糖脂的分解代謝在溶酶體水解酶的催化下進行。糖鏈的降解由各種外切糖苷酶催化,從非還原(遊離)末端逐個切下糖基。神經節苷脂的唾液酸基雖多在糖鏈的非還原末端,然而僅在無其他單糖作為末端基的情況下才能被唾液酸酶(又稱神經氨酸酶)催化水解。糖鏈完全水解後產生的神經酰胺有兩條去路:或作為合成的原料被利用,或進一步在神經酰胺酶的催化下水解為脂肪酸及鞘氨醇。硫酸鞘糖脂的分解還需要硫酸酯酶參加。
生物學作用 甘油糖脂主要存在於植物的光合組織及微生物的質膜中。在哺乳動物僅見於神經組織、睪丸和精細胞。生物學作用尚不詳。微生物質膜中的甘油糖脂結構差別很大,在微生物分類上有一定價值。植物中的甘油糖脂與葉綠素的構造及光合作用有關。
動物的神經組織中糖脂含量豐富。在中樞神經系統,腦苷脂(主要是半乳糖腦苷脂)的含量在腦白質中很高,此外鞘氨醇半乳糖苷及硫酸化半乳糖腦苷脂分佈亦很普遍,並有痕量甘油糖脂。神經節苷脂在腦灰質中含量較高。腦苷脂和硫酸腦苷脂還與鞘磷脂共同構成神經髓鞘的脂類。其合成不足或代謝障礙可嚴重影響髓鞘的發生和形成,破壞髓鞘的正常結構,引起病變。
鞘糖脂作為各種細胞質膜及細胞內膜不可缺少的成分,含量雖少而生物學作用相當復雜。糖脂作為膜的結構成分可改變膜的物理性質,如賦予質膜一定的剛性,使膜處於最適於發揮其作用的物理狀態。由於糖脂具有抗原性,質膜中的糖脂可作為表面抗原。例如,ABO(H)、Ii、Lea/ Leb等血型抗原決定簇不僅存在於紅細胞及其他非紅細胞質膜的糖蛋白糖鏈的末端,亦存在於質膜鞘糖脂糖鏈的末端。在同一組織中合成的糖蛋白及糖脂的寡糖鏈,其核心部分(還原末端)雖不相同,而末端部分(遊離的非還原末端)卻極其相似,因此可以共有某些功能。發生惡性轉化的細胞表面常出現腫瘤相關的糖脂抗原。例如,人、小鼠及倉鼠的黑素瘤細胞有共同的表面抗原CM3,而正常黑素細胞及其他腫瘤細胞基本不具有這種表面抗原。另外,在各種自發及誘發的人及鼠類肝癌細胞表面,GD3含量大幅度增高,並與腫瘤的發展相平行。制備腫瘤相關抗原的抗體對腫瘤的診斷及治療有一定意義。鞘糖脂還可作為細胞表面標志,對研究和認識免疫細胞、胚胎發育、細胞分化及腫瘤細胞很有意義。例如,人胸腺細胞可按其是否與識別 α半乳糖的花生凝集素 (PNA)相結合而分為PNA+PNA-兩個亞群。正常人周緣血中PNA+淋巴細 胞甚少(1~2%),急性淋巴性白血病及髓性白血病患者有半數出現PNA+細胞升高(≥20%);而慢性白血病及急性白血病的緩解期很少見PNA+淋巴細胞增高。因此測定周緣血中PNA+淋巴細胞的百分率有助於預後的判斷。質膜中的鞘糖脂與細胞表面生物活性分子,如受體及酶的關系,更是十分復雜。有的可能就是受體或受體復合物的一個組成部分,有的則為受體糖蛋白構象的維持者或功能的調節者。例如,GM1可作為霍亂毒素的受體,可高效、特異地中和霍亂毒素,抑制霍亂毒素的特異性毒性作用。具有 GM1寡糖結構的膜糖蛋白亦具有霍亂毒素受體的作用。此外,GD1b、GT1b可與破傷風毒素結合,並使之喪失毒性。破傷風毒素屬於神經毒。突觸膜中的上述神經節苷脂作為破傷風毒素的受體可能是產生破傷風神經癥狀的原因。巨噬細胞移動抑制因子 (MIF)的受體是一種巖藻糖化的腦苷脂。一些含糖肽類激素,如促甲狀腺素、黃體化激素、絨毛膜促性腺激素等,與其靶細胞的結合可被某些神經節苷脂抑制。幹擾素的抗病毒活性可被神經節苷脂增強,本來對幹擾素無反應的細胞經與神經節苷脂作用(外源性神經節苷脂嵌入於質膜中)後可變為有反應性。一些細胞外基質的細胞表面受體雖已鑒定為糖蛋白,但對一定的糖脂具有依賴性。它們在細胞表面共分佈,在體外共純化,去除糖脂則受體功能喪失。某些病毒亦與一定的鞘糖脂有親和性。以上種種事實都表明糖脂與多種生物活性分子乃至微生物的受體有密切關系。另外,位於神經末梢及突觸前膜的神經節苷脂及硫酸鞘糖脂,由於其所帶負電荷可與帶正電荷的乙酰膽堿、5-羥色胺、去甲腎上腺素及組胺等神經遞質結合,從而有助於神經沖動的傳導。除受體外,膜中酶的結構及功能亦與神經節苷脂有密切關系。例如,神經節苷脂GM1可活化Na+,K+-ATP酶,從而調節細胞膜的離子泵。腺苷酸環化酶及3',5'-環核苷酶磷酸二酯酶等亦可被一些神經節苷脂活化。此外,有一些膜蛋白並無疏水肽段,因此不能直接嵌入於膜脂雙層,它們借助於膜中的磷脂酰肌醇糖脂而“錨”定於膜上。例如乙酰膽堿脂酶、堿性磷酸酶、5'-核苷酸酶、氨基肽酶P以及脂蛋白脂肪酶等都以其羧基末端的氨基酸殘基與磷脂酰肌醇糖脂糖鏈末端借氨基乙醇共價結合。此外,腦及淋巴細胞表面的Thy-1抗原以及腫瘤細胞的癌胚抗原(CEA)等亦屬此例。
鞘糖脂參與細胞間的識別及粘合。例如,Gbos4Cer可能參與神經肌肉接頭的識別過程;GM2可能與視網膜頂蓋的識別過程有關。硫酸化鞘糖脂還參與細胞與細胞外基質的作用。一些細胞外基質糖蛋白,如層粘連蛋白、血小板反應蛋白及馮·維勒佈蘭德氏因子等,均可與硫酸化鞘糖脂結合。這種結合不但親和力高而且具有特異性。
鞘糖脂參與細胞的生長控制。鞘糖脂在質膜中的表達隨細胞周期而改變。細胞在體外培養時,細胞密度由於增殖逐漸增加,乃至彼此接觸。這時伴有一定種類的鞘糖脂合成增多,其在細胞表面的含量亦增高。這稱為“接觸延伸反應”。與此同時細胞延長,增殖減慢。加入外源性的相關鞘糖脂,可使旺盛增殖的細胞減慢增殖速度。可見,鞘糖脂合成的“接觸延伸反應”可能參與細胞生長的控制。轉化細胞及腫瘤細胞鞘糖脂的合成發生改變;某些糖脂的合成發生障礙,同時又可出現在對應的正常細胞很少或沒有的鞘糖脂,並有唾液酸化增強現象,還有一些鞘糖脂在細胞表面從隱蔽轉變為暴露。惡性細胞表面鞘糖脂的這些異常可能與其生長失控有關。分化誘導劑(丁酸鈉及維生素 A酸等)可增加某些糖基轉移酶的活性,促進一定的鞘糖脂合成,同時恢復某些轉化細胞在生長時的密度抑制(又稱接觸抑制)。有些神經節苷脂具有生長因子樣作用,另一些有生長抑制因子樣作用。
細胞表面的鞘糖脂在分化過程中亦發生顯著改變。在紅細胞和前腦細胞發育過程中以及成肌細胞和小腸上皮細胞分化過程中鞘糖脂的量或質發生變化。小腸上皮隱窩的未分化細胞表面含有乳糖基神經酰胺及葡萄糖腦苷脂,不含神經節苷脂GM3及其合成酶。在12小時內逐漸沿基膜向刷狀緣(絨毛頂)遷移,並進行分化,至分化為含消化酶的成熟細胞時上述兩種中性鞘糖脂的含量降至很低而GM3及其合成酶的含量顯著升高。此外,GM3還可促進肝癌細胞分化。
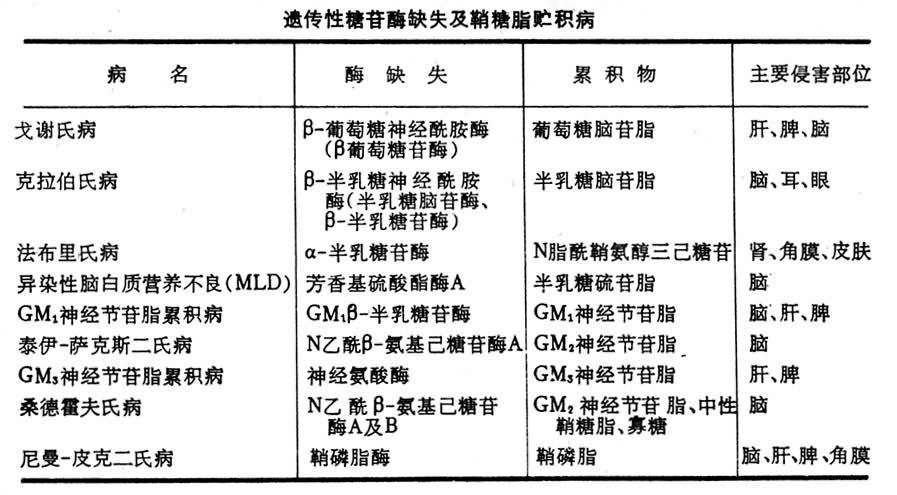
鞘糖脂貯積病 某一種或幾種糖苷酶的先天性缺失可導致一定種類的鞘糖脂在神經系統及其他器官中堆積而產生結構及功能障礙,如精神遲鈍,運動障礙,肝、脾腫大,眼盲、耳聾等,常致早夭。上表中列出某些鞘糖脂貯積病。這些先天性酶缺失為常染色體隱性遺傳(見脂質貯積病)。
遺傳性糖苷酶缺失及鞘糖脂貯積病