真核細胞分裂的基本形式。又稱間接分裂或核分裂。在這種分裂過程中出現由許多紡錘絲構成的紡錘體,染色質集縮成棒狀的染色體。1882年W.弗勒明最先將此種分裂方式命名為有絲分裂。
通過有絲分裂,作為遺傳物質的去氧核糖核酸(DNA)得以準確地在細胞世代間相傳。通過有絲分裂和細胞分化才能實現組織發生和個體發育。致癌因數通過使有絲分裂的調控失常而導致腫瘤發發生。
細胞在進行有絲分裂時,核和胞質都發生形態上的變化,稱有絲分裂期。兩次有絲分裂之間稱為分裂間期。有絲分裂期和間期合起來稱細胞周期。有絲分裂的準備工作,例如DNA復制、組蛋白合成、微管蛋白合成、能量準備等都是在間期進行的。
動植物的許多組織細胞都進行有絲分裂,特別是胚胎發育時期細胞的有絲分裂旺盛。高等植物的根尖和莖端分生組織,高等動物的骨髓和一些上皮組織,都是觀察有絲分裂現象的適宜部位。
過程 有絲分裂是一個連續的過程,為瞭描述方便起見,習慣上按先後順序劃分為前期、中期、後期和末期四個時期,在前期和中期之間有時還劃分出一個前中期。
前期 自分裂期開始到核膜解體為止的時期。間期細胞進入有絲分裂前期時,核的體積增大染色質構成的細染色線逐漸縮短變粗,形成染色體。因為染色體在間期中已經復制,所以每條染色體由兩條染色單體組成。核仁在前期的後半漸漸消失。在前期末核膜破裂,於是染色體散於細胞質中。動物細胞有絲分裂前期時靠近核膜有兩個中心體。每個中心體由一對中心粒和圍繞它們的中心粒周圍物質所組成。由中心體放射出星體絲,即放射狀微管。帶有星體絲的兩個中心體逐漸分開,移向相對的兩極(圖1)。這種分開過程推測是由於兩個中心體之間的星體絲微管相互作用,更快地增長,結果把兩個中心體推向兩極,而於核膜破裂後終於形成兩極之間的紡錘體。
前中期 自核膜破裂起到染色體排列在赤道面上為止。核膜的斷片殘留於細胞質中,與內質網不易區別,在紡錘體的周圍有時可以看到它們。
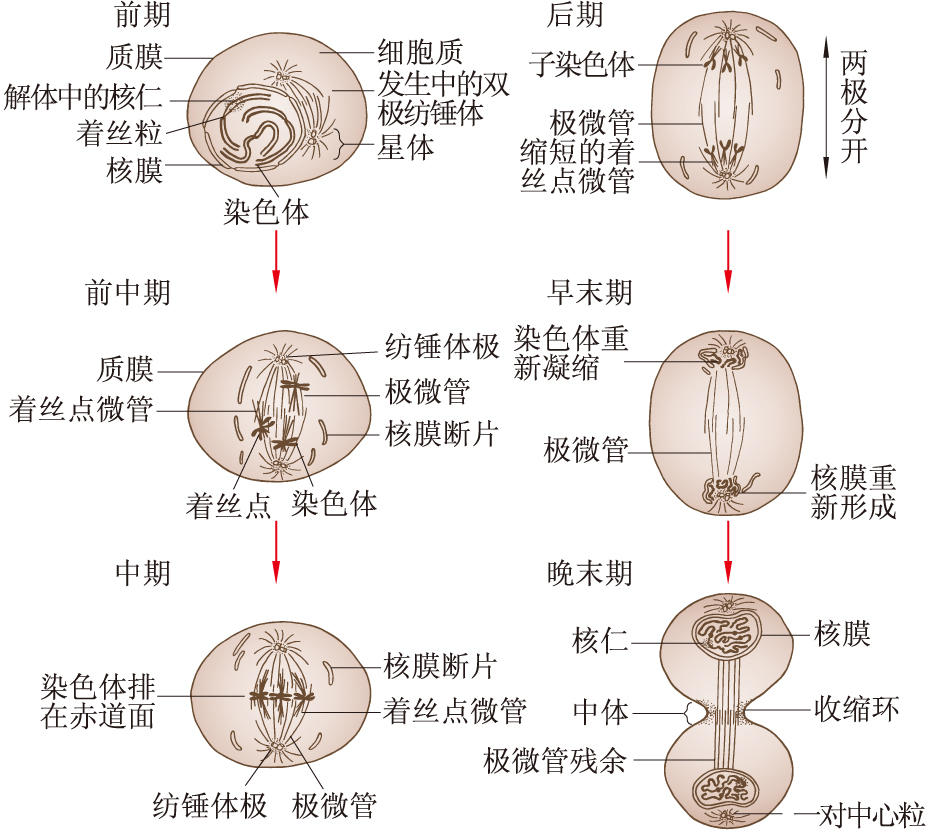 圖1 有絲分裂過程圖解
圖1 有絲分裂過程圖解
前中期的主要過程是紡錘體的最終形成和染色體向赤道面的運動。紡錘體有兩種類型:一為有星紡錘體,即兩極各有一個以一對中心粒為核心的星體,見於絕大多數動物細胞和某些低等植物細胞。一為無星紡錘體,兩極無星體,見於高等植物細胞(圖2)。
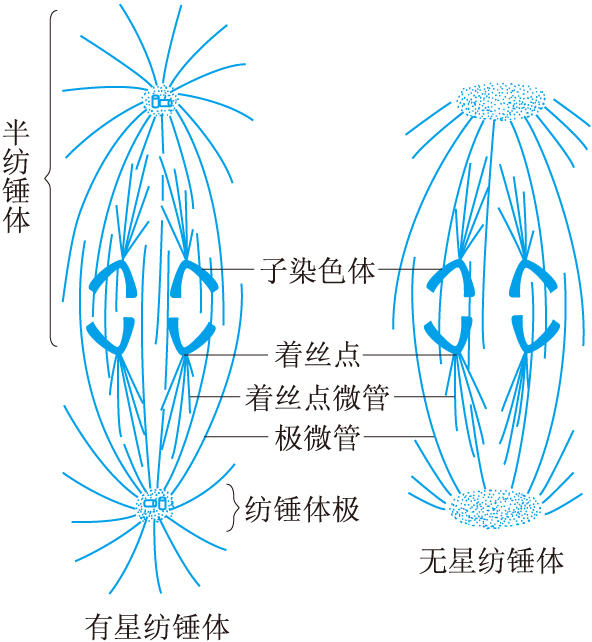 圖2 有星與無星紡錘體
圖2 有星與無星紡錘體
曾經認為有星紡錘體含有三種紡錘絲,即三種微管。一是星體微管,由星體散射出的微管。二是極微管,是由兩極分別向相對一級方向伸展的微管,在赤道區來自兩極的極微管互相重疊。現在認為極微管可能是由星體微管伸長形成的。三是著絲點微管,與著絲點聯結的微管,又稱著絲點絲或牽引絲。著絲點又稱動粒,是在染色體的著絲粒的兩側發育出的結構。有報告說著絲點有使微管蛋白聚合成微管的功能。無星紡錘體隻有極微管與著絲點微管。
核膜破裂後染色體分散於細胞質中。每條染色體的兩條染色單體的著絲點分別通過著絲微管與兩極相連。由於極微管和著絲微管之間的相互作用,染色體向赤道面運動。最後各種力達到平衡,染色體乃排列到赤道面上。
中期 從染色體排列到赤道面上,到它們的染色單體開始分向兩極之間,這段時間稱為中期。有時把前中期也包括在中期之內。中期染色體在赤道面形成所謂赤道板。從一端觀察可見這些染色體在赤道面呈放射狀排列,這時它們不是靜止不動的,而是處於不斷擺動的狀態。中期染色體濃縮變粗,顯示出該物種所特有的數目和形態。因此有絲分裂中期適於做染色體的形態、結構和數目的研究,適於核型分析。
後期 每條染色體的兩條姊妹染色單體分開並移向兩極的時期。分開的染色體稱為子染色體。子染色體到達兩極時後期結束。染色單體的分開常從著絲點處開始,然後兩個染色單體的臂逐漸分開。當它們完全分開後就向它們分別相對的兩極移動。這種移動的速度依細胞種類而異,大體上在0.2~5微米/分之間。平均速度為1微米/分。同一細胞內的各條染色體都差不多以同樣速度同步地移向兩極。子染色體向兩極的移動是靠紡錘體的活動實現的。
末期 從子染色體到達兩極開始至形成兩個子細胞為止稱為末期。此期的主要過程是子核的形成和細胞體的分裂。子核的形成大體上是經歷一個與前期相反的過程。到達兩極的子染色體首先解螺旋而輪廓消失,全部子染色體構成一個大染色質塊,在其周圍集合核膜成分,融合而形成子核的核膜,隨著子細胞核的重新組成,核內出現核仁。核仁的形成與特定染色體上的核仁組織區的活動有關。
細胞體的分裂稱胞質分裂。動物和某些低等植物細胞的胞質分裂是以縊縮或起溝的方式完成的。縊縮的動力一般推測是由於赤道區的細胞質周邊的微絲收縮的結果。微絲的緊縮使細胞在此區域產生縊縮,縊縮逐漸加深使細胞體最後一分為二。
高等植物細胞的胞質分裂是靠細胞板的形成。在末期,紡錘絲首先在靠近兩極處解體消失,但中間區的紡錘絲保留下來,並且微管增加數量,向周圍擴展,形成桶狀結構,稱成膜體。與形成成膜體的同時,來自內質網和高爾基器的一些小泡和顆粒成分被運輸到赤道區,它們經過改組融合而參加細胞板的形成。細胞板逐漸擴展到原來的細胞壁乃把細胞質一分為二(圖3)。細胞板由兩層薄膜組成,兩層薄膜之間積累果膠質,發育成胞間層,兩側的薄膜積累纖維素,各自發育成子細胞的初生壁。
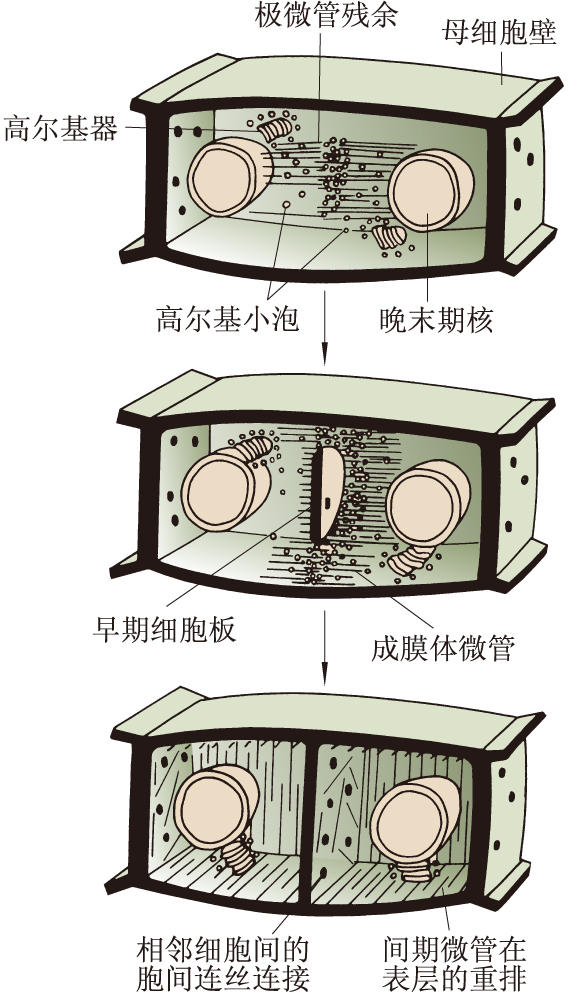 圖3 高等植物細胞體分裂過程略圖
圖3 高等植物細胞體分裂過程略圖
機制 染色體的集縮 構成染色體的細線在分裂前期縮短變粗,染色體的這種集縮運動是通過染色線的螺旋化實現的。染色質濃縮過程和細胞質中的某些因素有關。如果用實驗方法使分裂期細胞與間期細胞融合,可以觀察到間期細胞染色質會提前集縮成染色體。已經證明是M期細胞中產生的MPF因子促使染色體集縮。
紡錘體的形成 由微管蛋白聚合成紡錘體微管的過程。微管蛋白的聚合有兩種基本形式:一種是自我裝配型,另一種是位點起始裝配型,後者有特殊位點作為聚合的起始部位,前者沒有這種特殊位點。形成紡錘體時的位點統稱為“微管組織中心”(MTOC)。中心體和著絲點都是MTOC,它們在離體情況下都能表現出使微管蛋白聚合成微管的能力。紡錘體的形成顯然和這些MTOC的活動是分不開的。
中期染色體運動 用藥物(秋水仙素、巰基乙醇等)破壞紡錘體,則染色體不能排列到赤道面,除去藥物後,紡錘體重新形成,則染色體又能排列到赤道面,由此可見,染色體向赤道面的排列和紡錘體的活動有關。由輻射損傷或其他原因造成的沒有著絲點的染色體斷片不能排列到赤道面上。因此說明,染色體向赤道面的排列和著絲點的活動有關。用微束紫外線照射減數分裂時二價體的一側著絲點或著絲點絲,則染色體不能定位於赤道面,而偏近於未受照射的著絲點所面向的一極。這說明染色體在赤道面的定位必須兩個著絲點及與兩極相連的兩側著絲點絲都正常地發揮作用。根據以上事實和其他觀察,推測在前中期時兩個著絲點分別以著絲點絲與兩極相連,靠兩極牽引力的平衡,使染色體定位於赤道面上。除這種牽引平衡的力量外,還可能有其他一些因素起輔助作用。
後期染色體運動 後期時兩組子染色體向兩極移動,而在有些細胞兩極也被推開更遠。關於這種運動的機制尚無定論。後期時著絲點微管在向極的末端不斷解聚,因而逐漸變短。這可能是使染色體被拉向兩極的重要原因。因為在體外實驗中給模型細胞添加D2O以阻抑微管的解聚時,則染色體向兩極移動過程停止,反之,如果添加少量秋水仙素以促使微管解聚速度加快,則染色體向兩極移動速度也加快。有些細胞在分裂後期兩極分開更遠可能是由下述機制造成的:來自兩極的極微管在赤道區互相重疊,微管蛋白在它們的自由末端聚合而使微管加長。這些重疊的來自兩極的微管互相滑動,使兩極推開更遠(圖4)。滑動的動力可能是由類動力蛋白ATP酶所產生的。這種分子使ATP水解,釋放出滑動所需的能量。這種推測的一個依據是dynein的許多抑制劑,都可阻抑分裂後期的兩極分開過程。
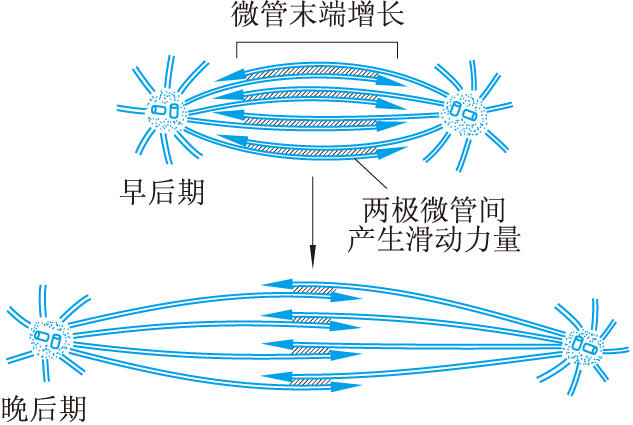 圖4 分裂後期兩極分離更遠的機制示意圖
圖4 分裂後期兩極分離更遠的機制示意圖
與兩極的相互遠離過程不同,染色體向兩極移動過程受dynein抑制劑的影響不顯著。推測染色體向兩極移動所需能量可能是靠另外一種尚不明瞭的機制產生的。
持續時間 有絲分裂期所用時間,因生物和細胞的種類不同而有很大差別。一般要1~3小時,但快者有10多分鐘就可完成,慢者需5~6個小時,甚至更長時間。通常,胚胎發生早期細胞分裂快,而成體組織細胞分裂慢。
在有絲分裂期中通常是前期和末期時間較長,而中期和後期時間較短。尤其是後期一般需要時間最少。
控制 許多理化因素都影響有絲分裂過程。對於植物和低等動物的細胞,在一定范圍內(8~25℃)溫度越低,細胞周期越長,有絲分裂過程也越慢。接近零度的低溫可抑制紡錘體的形成,使細胞受阻在有絲分裂中期。電離輻射,如X射線和γ射線能抑制細胞進入有絲分裂。許多化學藥物都有抑制有絲分裂的作用,如秋水仙素、對二氯苯、巰基乙醇、咖啡堿等。促進有絲分裂的化學物質有植物血球凝集素(PHA)、鯡精胺、激動素、赤黴素等。
在機體內環腺苷酸(cAMP)增加時抑制有絲分裂,而環鳥苷酸(cGMP)增加時促進細胞有絲分裂。