免疫系統對一種或多種抗原的特異無反應狀態。表現為對該抗原的刺激既不產生抗體,也不產生細胞免疫。這種無反應狀態和有反應狀態一樣也是有免疫學特異性的,也就是說對某種抗原耐受的機體,對其他抗原仍能照常產生免疫反應。又有完全耐受和不完全耐受之分,前者表現為對耐受抗原完全無反應;而後者則為對抗原的反應微弱,或隻能產生少量低親和力的抗體如IgM。正常機體的免疫系統對外來的抗原應表現為正反應,也就是產生抗體或細胞免疫,最終把不是自身成分的外來抗原清除。正常的免疫系統對自自身成分則應表現為耐受狀態,不發生免疫反應,不排斥自身的組織或細胞。免疫系統對自己和非己抗原成分的這兩種截然不同的反應方式對種系發生或個體發生都十分重要。沒有對自身抗原的免疫耐受,從種屬到個體都不可能存在,在發育成長之前任何個體就被自身免疫系統所消滅。沒有對外來抗原的特異免疫反應,特定的生物即不能形成,也不可能在多種外源生物侵襲下成長壯大。自身耐受的維持也是個體生存、正常生理功能得以進行的保證。反之自身耐受的破裂產生自身免疫性疾病。此外,過敏性疾病的阻止和控制,器官移植或骨髓移植均涉及免疫耐受問題。
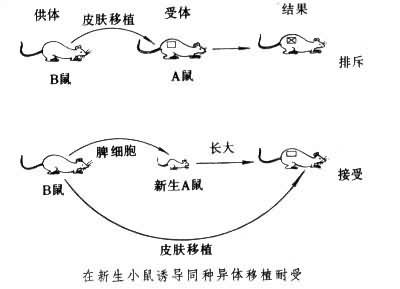
兩種免疫耐受現象 1945年美國R.D.歐文觀察到異卵雙生小牛的胎盤上有共同的血管供應。在雙生小牛中每一頭牛身體內有它自身的和同胎另一頭小牛的兩種血型的紅細胞。他認為這種一個個體之所以能耐受另一個體不同血型的細胞的存在,是由於胚胎期雙生小牛在子宮內有共同的血管供應,它們的血細胞在胚胎期在兩者體內交互流通。他進一步觀察發現這對異卵雙胎小牛可耐受彼此相互的皮膚移植,這種現象稱為天然耐受。在此基礎上,P.B.梅達沃制成實驗耐受模型,並因此獲1960年諾貝爾生理學或醫學獎。他用遺傳基因不同的小鼠A和B做實驗,把B鼠的脾細胞給新生出的A小鼠註射,待A鼠成年之後,就可接受移植自B鼠的皮膚。而對來自其他品系的皮膚移植物仍有正常排斥反應(見圖)。原因是胚胎期或新生期(如新生A小鼠)接受某種抗原(如B鼠的脾細胞),待成年之後,它的免疫系統則視B鼠的組織或其他抗原為自身的而不予排斥。而對除 B鼠抗原以外的異體成分仍能識別和排斥。梅達沃又給已產生耐受的 A成年鼠註射正常成年同基因A鼠的淋巴細胞,則它對B鼠皮膚的耐受就被打破,這一重要的發現說明A鼠在胚胎期受到B鼠抗原刺激,能與B系抗原起反應的淋巴細胞克隆便被抑制或排除,於是A鼠對B鼠抗原產生耐受,當這種已有耐受性的A鼠接受正常A鼠的淋巴細胞註射,體內又重新出現各種有功能的淋巴細胞,於是又出現對B系抗原的排斥反應。這說明註射進來的正常A鼠淋巴細胞可以打破耐受,也就是耐受A鼠體內的淋巴細胞的被抑制或排除是造成耐受的原因,這一點把人們對免疫耐受的認識從整體水平進一步推進到細胞水平。應用結構更清楚的蛋白質抗原誘導人工耐受,更能深入瞭解誘導耐受產生的分子結構。
影響免疫耐受的因素 從上述實驗研究可知,免疫耐受是免疫系統與特異抗原在特定條件下相互作用的結果,所以耐受的產生主要與機體免疫系統狀態和抗原的類型、數量有關。免疫系統在不成熟的胚胎期或新生期較易產生耐受,而誘導成年動物的免疫耐受隻有在免疫系統功能低下或受抑制(如全身X射線照射,服用免疫抑制藥物如環磷酰胺、硫唑嘌呤等,以非特異地破壞淋巴細胞)時才有可能。機體的遺傳背景也影響耐受現象的產生,有自身免疫或過敏反應傾向的機體難以誘導耐受。影響產生免疫耐受的抗原條件是抗原的類型,以卡介苗(BCG)為例,可溶性形式的BCG容易產生耐受,而聚合形式的則是免疫原,溶解狀態的單體形式是耐受原而聚合的沉淀物就是很好的免疫原。抗原的劑量對產生耐受也很重要,特別大量和特別小量的抗原易於誘導耐受。不易代謝的物質是耐受原,如肺炎鏈球菌多糖,人工合成的D-多肽不易被酶降解,在體內雖可被吞噬細胞吞噬,但不易被消化,可不斷釋放到循環中,與抗體結合後就不易激發免疫反應而導致免疫耐受。
免疫耐受的機理 免疫反應過程由單核巨噬細胞、T細胞和B細胞的分工和協作完成,所以免疫耐受過程亦與三類免疫細胞的功能有密切關系。單核吞噬細胞處理-呈遞抗原給T和B細胞,是引起免疫反應的起始細胞,單核吞噬細胞的先天發育障礙或後天因素造成的缺損都可誘生免疫耐受。實驗證明新生期容易產生耐受的原因亦與此時單核吞噬細胞數量較少、功能不成熟有關。T和B細胞在產生免疫耐受中的作用尚知之不詳。總的來說,T和B細胞都可誘導耐受的產生,T細胞較B細胞更易誘導。T細胞產生耐受的潛伏期短,持續時間長,而B細胞建立的耐受潛伏期長,持續時間短,這與它們個體發生過程的不同有關。T細胞在胸腺內分化發育,而胸腺造就T細胞的能力在胚胎期和新生期最活躍,到青春期後胸腺逐漸退化萎縮。所以成年動物的T細胞很少有更新,因此形成耐受的T細胞就能持續很長時間。反之,B細胞在骨髓成熟,而骨髓終生有功能,不斷有新的B細胞投入運作,耐受的B細胞不斷被新的細胞所代替,因此B細胞的耐受狀態持續時間短。
早期研究證明,在胚胎期與自身成分起反應的細胞克隆被排除就產生自身耐受,但新的資料證明正常機體內存在著針對自身某些成分起反應的淋巴細胞克隆,隻是在正常情況下處於抑制狀態,不能活化起反應而已。在異常情況如感染、電離輻射中就可活化產生自身抗體。因此在免疫耐受中除細胞和因子的作用外,免疫調節機制也有重要作用。
有關B細胞產生耐受的學說,最早有著名的克隆流產學說:機體內存在著成千上萬種能與體內(自己的)和外界非己的抗原起反應的細胞克隆。每一種細胞克隆專一地與某種抗原起反應,這就產生瞭免疫反應的特異性。若免疫系統在不成熟時即與某種抗原(自身的或異體的)相遇,這就可使針對這種抗原起反應的淋巴細胞克隆夭折(流產),待到成年之後機體再與這個抗原相遇也不會發生反應。而體內其他的無數種細胞克隆繼續存活。成年後,某種抗原出現即有特異的細胞與它起反應,產生抗體或細胞免疫。新的證據證實,B細胞在體內發育的早期階段,細胞膜表面帶有IgM受體,此時與抗原相遇則表現為耐受。B細胞在體內進一步發育成熟則細胞膜表面既帶有IgM又帶有IgD受體,此時與抗原相遇則表現為免疫反應。還有人證明,成年鼠的脾細胞中沒有帶針對自身抗原的受體,而將同一鼠的骨髓細胞取出,讓它在體外成熟為B細胞,則其細胞表面就出現能與自身抗原起反應的受體,這說明B細胞在體內由骨髓內前體細胞發育成熟的過程中,與體內抗原相遇則該細胞的克隆被排除。若該骨髓前體細胞在體外發育成熟,不與自身抗原接觸,則其細胞克隆不被排除,卻能存活下去。再遇自身抗原就能起反應,而不表現為耐受。
有人認為T細胞在免疫耐受產生中的作用似B細胞,也有特異的自身反應性T細胞克隆的流產或排除,從而導致耐受。1970年格爾雄發現能抑制 B細胞產生抗體的抑制性T細胞 (Ts)。後來單克隆抗體技術又證明人類的Ts表面的標志是CD8分子(小鼠的是Lyt.2+)。實驗證明這種Ts細胞不但使B細胞反應抑制,也可抑制T細胞的功能。對某一抗原有耐受性的小鼠體內就有數量多、功能強的Ts細胞,把耐受小鼠的脾細胞註射到正常小鼠體內,則使正常小鼠也不能對這種抗原起反應,而接受瞭耐受動物的Ts細胞的動物,也表現為免疫耐受狀態。這種被動轉移Ts細胞的實驗說明Ts細胞在免疫耐受狀態的產生方面也是重要的。
免疫網絡學說 認為抗淋巴細胞的抗體可以與淋巴細胞的抗原受體結合而阻止其分化發育,另一方面抗獨特型抗體可以激活Ts細胞而誘導產生免疫耐受。綜上所述免疫耐受現象是由抗原誘導產生的特異無反應狀態,參與這一過程的細胞、分子甚至某些基因成分是很復雜的,而耐受狀態的維持及解除又有多種體內、體外的因素參加。
免疫耐受現象的臨床意義 這方面的研究主要用以解釋自身免疫性疾病的發病機理,維持自身耐受以阻止或治療自身免疫病,以及應用誘導耐受的手段來維持移植器官的存活,阻止排斥發生。免疫耐受狀態的維持必須有耐受原持續存在,自身抗原不僅在胚胎期誘導免疫系統對它的耐受,而且成年以後仍不斷地刺激新生的淋巴細胞來維持耐受。所以通常情況下,機體的自身耐受十分穩定。在異常情況下,如免疫細胞自發突變,受抑制的細胞又發生活化,免疫抑制藥物破壞Ts細胞,則抑制自身抗體產生的能力可下降或消失。某些病原生物與人體某些組織有共同的抗原結構。當乙型溶血性鏈球菌感染時活化的TH細胞輔助B細胞產生抗體,由於乙型溶血性鏈球菌與人體的腎小球基底膜之間有共同的抗原結構,這打破瞭機體對腎小球基底膜的耐受而引起腎小球腎炎。已有報告在成年大鼠、狗、猴和人體內誘導建立對同種異體移植物的免疫耐受。在人類骨髓的同種異體移植已有成功的報告,但迄今對整個移植器官的耐受還沒有成功。在對異體移植器官建立免疫耐受的嘗試中,人們在移植物方面尋找抗原性匹配,供者與受者之間抗原越相同或相近則越易耐受。從近親、父母兄弟姐妹中尋找供體移植成功的可能性越大。另一方面還需從移植物中盡量去除能引起受體免疫反應的成分。在受體方面也須盡量排除能與移植物發生免疫反應的細胞(如TH細胞或活化的T細胞),並且在受者體內設法活化或誘導天然抑制細胞。應在受者體內建立對移植物的免疫耐受,而不是依賴免疫抑制藥物來維持移植器官的存活。