植物葉綠體的類囊體膜或光合細菌的載色體利用光能從腺苷二磷酸(ADP)和無機磷酸(Pi)合成腺苷三磷酸(ATP) 的過程。1954年D.I.阿爾農等和A.W.弗倫克爾先後在植物葉綠體和光合細菌載色體中發現此反應。光合磷酸化在光合作用能量轉換中起關鍵作用。將由光能推動形成的電位能轉變為ATP分子中的化學能,貯存並推動碳同化等反應。
類型 按偶聯磷酸化的光合電子傳遞鏈的不同,光合磷酸化可分3種類型。
非循環(非環式)光合磷酸化 ATP形成偶聯於電子從H2O到NAD+的需光系統Ⅱ(PSⅡ)和光系統Ⅰ(PSⅠ)參與的非循環電子傳遞。在這個途徑中,水分解放出氧氣,其電子經一系列電子載體傳遞,最後將NAD+(Eò=-0.34V)還原。電子傳遞過程中偶聯形成ATP。總反應式為:

循環(環式)光合磷酸化 ATP 形成偶聯於隻牽涉PSI、不釋放氧氣也不氧化或還原其他物質的循環電子傳遞。用下式表示:
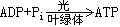
循環與非循環光合磷酸化二者可能共用一部分電子傳遞鏈(見光合電子傳遞)。
假循環(假環式)光合磷酸化 離體葉綠體在不外加電子受體時照光,可慢慢地吸收氧氣。加入天然電子受體Fd則氧氣的吸收增加,因為Fd被PSI還原,還原態Fd與O2反應產生過氧化基O3-,O3-再被還原態Fd還原為H2O2。
Fd(還原態)+O2→O2+Fd(氧化態)
Fd(還原態)+O2+2H+→H2O2+Fd(氧化態)
在Fd所催化的體內假循環電子傳遞中,最終電子受體是氧,同時ATP相伴形成。
與二氧化碳同化的關系 在三碳植物的光合作用中,同化1分子CO2並還原到[CH2O](碳水化合物)水平,需2分子NADPH和3分子ATP,其ATP/2e-比值應為1.5。非循環電子傳遞鏈從 H2O傳遞4個電子到 NADP+形成2分子NADPH,並可形成3分子ATP,即ATP/2e-比為1.5。
在四碳植物的四碳途徑中,每同化1分子CO2需要5分子ATP,2 分子NADPH,較一般三碳植物的光合碳循環多需要2分子ATP。葉肉細胞葉綠體雙羧酸途徑的這種“額外”需要,由維管束鞘細胞的無基粒葉綠體所進行的循環光合磷酸化來提供。
循環光合磷酸化產生的 ATP,還可作為蛋白質合成、葡萄糖同化、K+吸收與氣孔運動等的能源。它在光合作用的演化過程中出現較早。
與電子傳遞的偶聯 多年來人們探尋連接電子傳遞和磷酸化之間的高能中間物質(或狀態)的存在及其性質。沈允鋼和沈鞏楙於1962年,G.欣德和A.T.賈根多夫於1963年,先後發現小麥和菠菜葉綠體在光下形成某種高能態,在黑暗中可推動ADP和Pi形成ATP。以後發現,不同植物的葉綠體和光合細菌載色體照光後均可形成高能態,表明它的存在是普遍的。從這種高能態與光合磷酸化之間的關系判斷,它就是光合磷酸化的中間物。
在葉綠體、線粒體和光合細菌載色體中,進行光合磷酸化或氧化磷酸化時,電子傳遞和ATP形成之間的偶聯機理是相似的。對這個偶聯機理的解釋有化學假說、化學滲透假說、膜上區域化質子假說和變構假說。
化學假說 英國E.C.斯萊特於1953年提出,假設在偶聯過程中形成高能中間物(~X),其能量足以推動ATP形成。這個假說提出之後,始終未能分離或證實所假設的高能中間物。
化學滲透假說 1961年英國P.D.米切爾提出,認為不是中間化合物,而是以膜兩側的電位差和離子(質子)濃度差推動 ATP的形成。從葉綠體、線粒體和載色體等不同能量轉換系統中得到的證據,使此學說成為生物力能學的基本理論,米切爾因而獲得1978年諾貝爾化學獎。這個假說有4個基本的前提或假設:①葉綠體或線粒體偶聯磷酸化的膜(類囊體膜、線粒體內膜)主要由疏水的脂肪及多肽所組成,H+不易透過;②有一個定向轉移質子的氧化還原系統嵌在膜上,由其中的氫離子載體及電子載體從膜的一側水相中吸收一個質子,在膜的另一側放出一個質子,從而形成跨膜的質子濃度差或△pH。這種機構也稱為質子泵。膜兩側的質子濃度差還引起一個膜兩側的電位差(△ψ)。△pH 和 △ψ二者共同組成質子動子勢(pmf),是推動ATP形成所需的高能態;③膜上有一個偶聯於質子轉移的陰陽離子交換擴散系統,離子交換可以控制△ψ 和△pH在pmf中所占的比重;④膜上嵌有一轉移質子的可逆的ATP酶系統,即加水/去水系統。由氧化還原反應所產生的質子,在該系統作用下,使ADP與Pi脫水形成ATP。質子返回時,轉移數與ATP形成量之間有準量關系。
大量實驗結果支持米切爾的設想。當對無緩沖的葉綠體懸浮液照光時,引起H+吸收,同時類囊體內水相變酸,類囊體膜內外質子濃度差可達3~3.5pH單位,膜電位差可達10~100毫伏。進行光合磷酸化時,質子梯度減少,並加速瞭反映類囊體內膜電位差形成(<20納秒)的515納米處吸收的衰減,即降低瞭pmf。△pH和△ψ 在光合磷酸化反應起始或恒態下的貢獻不同,它們可以單獨或共同推動ATP形成。
化學滲透假說並不排除化學假說所假設的有~X 可推動離子泵和ATP形成的可能性;也不排除膜上的電子傳遞系統和ATP酶系統之間有更直接的能量轉換方式。
膜上區域化質子假說 R.J.P.威廉斯1961年提出,假定與氧化還原反應同時產生的質子,在膜的疏水區域內積累。這些無水質子在膜內以比在水中高得多的速度沿一系列氫鏈傳遞到達ATP酶,推動 ATP和Pi脫水形成ATP。該假說強調區域化質子的化學勢和電子傳遞鏈上蛋白質-電子載體的構象變化在能量轉換中起決定性作用,跨膜的△pH和△ψ 勢差不直接參與能量的積聚、貯存和轉換。
變構假說 P.D.博耶在1965年提出,認為能量轉換的中間步驟不是“高能化合物”或者跨膜的質子濃度差,而是膜上有關的蛋白質的構象變化。這個假說在1973年以後發展為:從氧化還原反應的能量以蛋白質構象變化的形式提供,使在催化部位早已形成的非共價結合的ATP釋放,而ATP的形成則不需要能量。
偶聯因子H+-ATP酶復合體的結構與功能 光合磷酸化反應最終形成 ATP的關鍵酶是ATP酶復合體(CF1-CF0)。起催化作用的主要是此復合體中的親水性蛋白偶聯因子(CF1)。CF1位於葉綠體未垛疊部分的類囊體膜的外表面上,為直徑10納米的顆粒,可以用乙二胺四乙酸(EDTA)從膜上洗脫,經活化處理後,可表現出催化ATP水解的ATP酶活力。CF1由α,β,γ,δ,ε等5種不同的亞單位組成,各亞單位都是多肽,它們分別承擔不同的催化或調節功能。CF0是嵌於膜上的疏水性蛋白,由4種不同的亞單位組成。其功能是作為膜的質子通道,將類囊體膜內部質子跨膜傳導給CF1。
參考書目
上海植物生理研究所、植物研究所:《光合作用研究進展》,科學出版社,北京,1977。
上海植物生理研究所、植物研究所:《光合作用研究進展》,第二集,科學出版社,北京,1980。
D.O.Hall and K.K.Rao,Photosynthesis,4th ed.,Edward Arnold Ltd.,New York,1987.