大腦、小腦、腦幹和脊髓血液迴圈的統稱。高等脊椎動物大腦迴圈在腦迴圈中所占的比重較大,所以也叫大腦迴圈,又因腦迴圈主要在顱腔內進行,故又叫顱內迴圈。腦迴圈是特殊區域迴圈的最重要組成部分。在進化過程中,隨著腦量的增加,腦功能更加複雜活躍,腦的能量消耗也大大增加,與之相適應的腦迴圈也更加複雜完善。例如人腦的耗氧量約為全身耗氧量的1/5,人腦血流量約占全部心輸出量的13~15%。充足的腦血流量是保證腦部正常活動的首要條件。腦血流供應不足很快會嚴重影響腦的功能。大腦腦皮層對腦循環缺血和血中缺氧非常敏感,腦循環血中缺氧半分鐘或完全阻斷腦血流10秒鐘即會導致昏迷,缺氧3分鐘可能造成腦神經細胞的不能恢復的損傷,缺氧6分鐘可以致死。由此可見,腦循環關系到動物的生死存亡。腦循環供給中樞神經系統營養並排除其有害的代謝產物,從而維持其正常功能。
結構 動脈的分佈 脊椎動物的腦血流來自兩對動脈,包括椎動脈和頸內動脈各1對(圖1)。
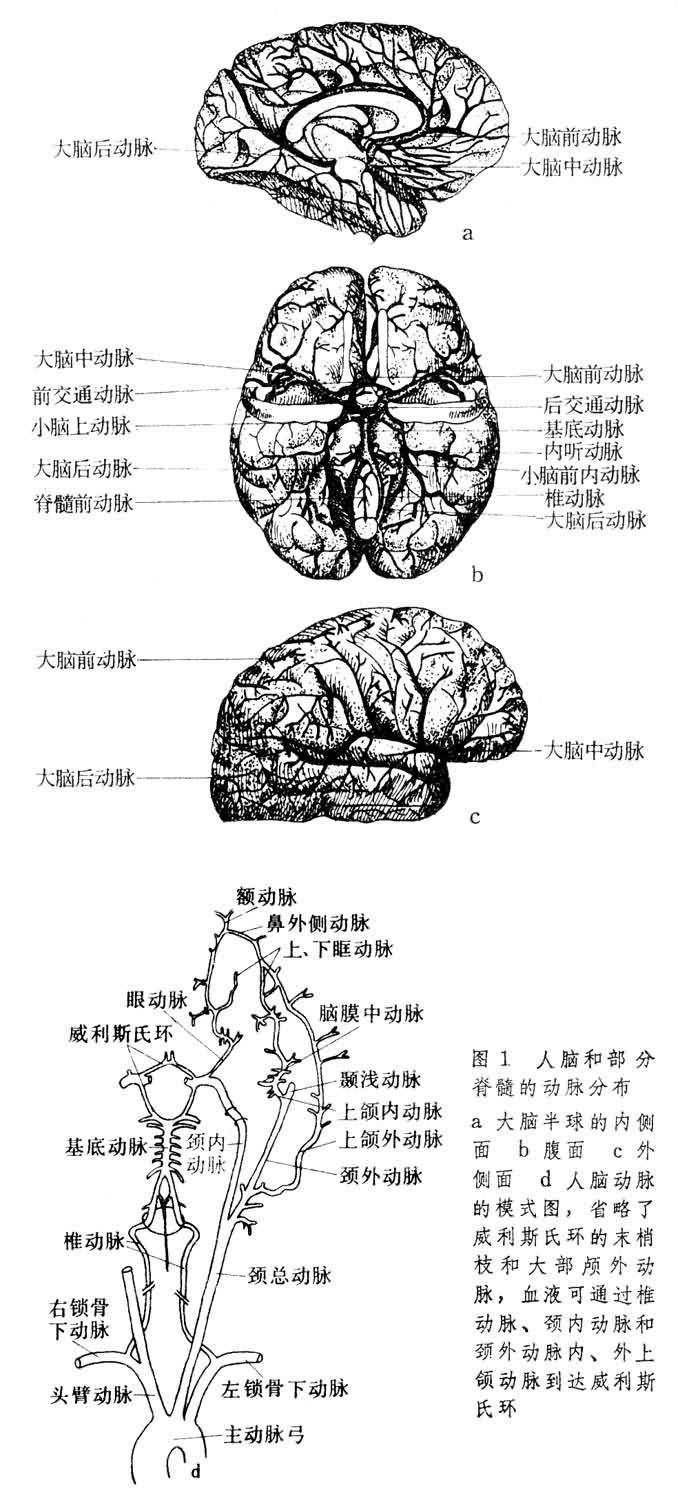 左、右椎動脈從枕骨大孔進入顱腔匯成基底動脈,然後與後交通動脈、頸內動脈、前交通動脈會合形成大腦動脈環,它首先由英國解剖學傢T.威利斯發現,所以又叫威利斯氏環,此環是腦血流分佈的集合管,它與基底動脈都屬腦循環的一級動脈。由此環發出6條大腦動脈供血給大腦、腦幹,從基底動脈發出1對到小腦的動脈,另外在椎動脈匯成基底動脈以前發出脊髓前動脈。頸內動脈供血到大腦半球兩側的前部和中部,椎動脈和基底動脈供血到小腦及大腦的枕葉和後窩。多數脊椎動物包括人類在內,椎動脈較細,供血量較少。頸內動脈較粗,供血量較多。少數脊椎動物椎動脈比頸內動脈粗,供血量較多。另外血液也可以通過頸外動脈的內、外上頜枝到達威利斯氏環。註入頸內動脈的血隻供同側的大腦半球。兩大腦半球之間沒有血管交叉。甚至威利斯氏環的吻合枝內也無大量血流通過。因此當一側頸內動脈阻塞時,常隻引起同側的腦缺血癥狀,特別在老人是如此。雖然大腦動脈的前毛細血管之間有吻合枝,但通過的血流量很少,常不能滿足所分佈流域腦組織的代謝需要。腦實質的動脈與身體其他部位類似大小的動脈相比,其組織結構的特點是肌層較薄,但彈性纖維較多,並有不同的排列形式。
左、右椎動脈從枕骨大孔進入顱腔匯成基底動脈,然後與後交通動脈、頸內動脈、前交通動脈會合形成大腦動脈環,它首先由英國解剖學傢T.威利斯發現,所以又叫威利斯氏環,此環是腦血流分佈的集合管,它與基底動脈都屬腦循環的一級動脈。由此環發出6條大腦動脈供血給大腦、腦幹,從基底動脈發出1對到小腦的動脈,另外在椎動脈匯成基底動脈以前發出脊髓前動脈。頸內動脈供血到大腦半球兩側的前部和中部,椎動脈和基底動脈供血到小腦及大腦的枕葉和後窩。多數脊椎動物包括人類在內,椎動脈較細,供血量較少。頸內動脈較粗,供血量較多。少數脊椎動物椎動脈比頸內動脈粗,供血量較多。另外血液也可以通過頸外動脈的內、外上頜枝到達威利斯氏環。註入頸內動脈的血隻供同側的大腦半球。兩大腦半球之間沒有血管交叉。甚至威利斯氏環的吻合枝內也無大量血流通過。因此當一側頸內動脈阻塞時,常隻引起同側的腦缺血癥狀,特別在老人是如此。雖然大腦動脈的前毛細血管之間有吻合枝,但通過的血流量很少,常不能滿足所分佈流域腦組織的代謝需要。腦實質的動脈與身體其他部位類似大小的動脈相比,其組織結構的特點是肌層較薄,但彈性纖維較多,並有不同的排列形式。
腦毛細血管 內皮細胞之間聯系緊密,沒有其他組織毛細血管那樣的孔或窗。而是形成一種無孔膜,內皮細胞之外有致密的基底膜包圍著,再外層則是星形膠質細胞的血管周足(終足)。大約有80%左右腦毛細血管表面被周足所包圍。這些結構形成層層壁壘,使得腦毛細血管中血液的很多物質特別是脂溶性較低的大分子量物質如蛋白質、抗菌素,某些大分子染料等難以通過,而隻允許大多數離子和葡萄糖等小分子物質通過,從而形成所謂的血腦屏障。
腦部靜脈血的回流路徑 首先經過上矢狀竇和直竇。直竇來源於下矢狀竇和蓋倫氏大靜脈。上矢狀竇和直竇在顱骨中縫處消失形成左右橫竇。橫竇的外側下方形成乙狀竇,然後穿過顱骨入頸內靜脈。在穿顱前乙狀竇還接受來自上、下巖竇的靜脈血。巖竇與海綿竇、竇狀靜脈叢、眶靜脈叢和面靜脈相連。海綿竇經基底靜脈叢與很多椎靜脈和靜脈竇相通(圖2)。
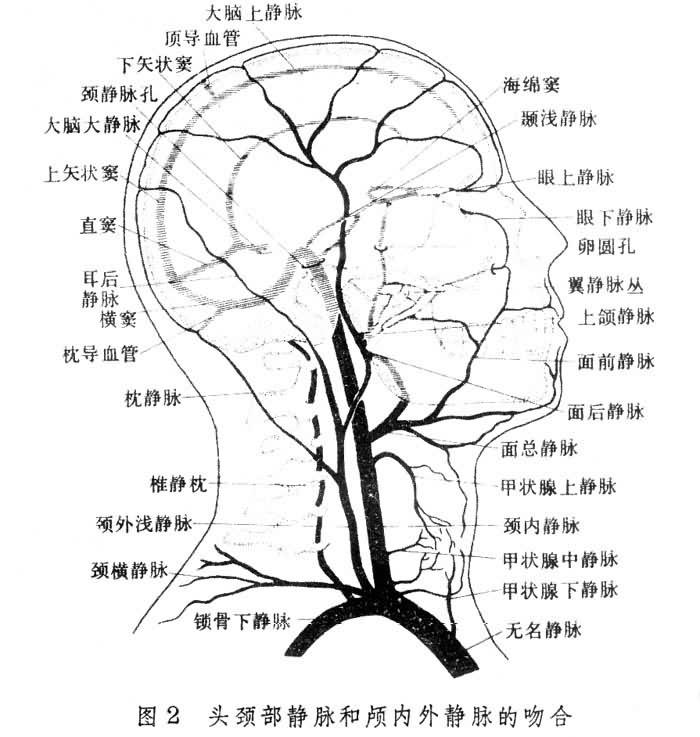 與腦動脈相比,腦靜脈管壁較薄。與身體其他部位的靜脈不同,腦靜脈管壁中沒有靜脈瓣,靜脈血的回流依賴高位的勢能。上述比較復雜的回流路徑可以歸納為:大部腦靜脈血經腦深部靜脈和腦血竇流入頸內靜脈;小部腦靜脈血經眼部翼狀靜脈叢,進入靜脈導血管再到頭皮,最後流入椎管中的椎旁靜脈系統。由圖2可見腦靜脈系統有大量交通枝靜脈叢,這些結構有重要的生理意義,即使兩側頸內靜脈都被阻塞,大腦靜脈血仍可經椎靜脈和頸外靜脈系統完成其回流。
與腦動脈相比,腦靜脈管壁較薄。與身體其他部位的靜脈不同,腦靜脈管壁中沒有靜脈瓣,靜脈血的回流依賴高位的勢能。上述比較復雜的回流路徑可以歸納為:大部腦靜脈血經腦深部靜脈和腦血竇流入頸內靜脈;小部腦靜脈血經眼部翼狀靜脈叢,進入靜脈導血管再到頭皮,最後流入椎管中的椎旁靜脈系統。由圖2可見腦靜脈系統有大量交通枝靜脈叢,這些結構有重要的生理意義,即使兩側頸內靜脈都被阻塞,大腦靜脈血仍可經椎靜脈和頸外靜脈系統完成其回流。
腦血管的神經支配 腦血管具有雙重神經支配,腦的動靜脈都有很多神經纖維包圍著。其中無髓鞘神經纖維傳導血管運動沖動;有髓鞘神經纖維屬感覺纖維,牽拉這些纖維會引起痛覺。頸內動脈和椎動脈的交感神經支配來自交感鏈的星狀神經節和上頸神經節。副交感神經纖維來自面神經,通過膝狀神經節和巖淺大神經(圖3)。
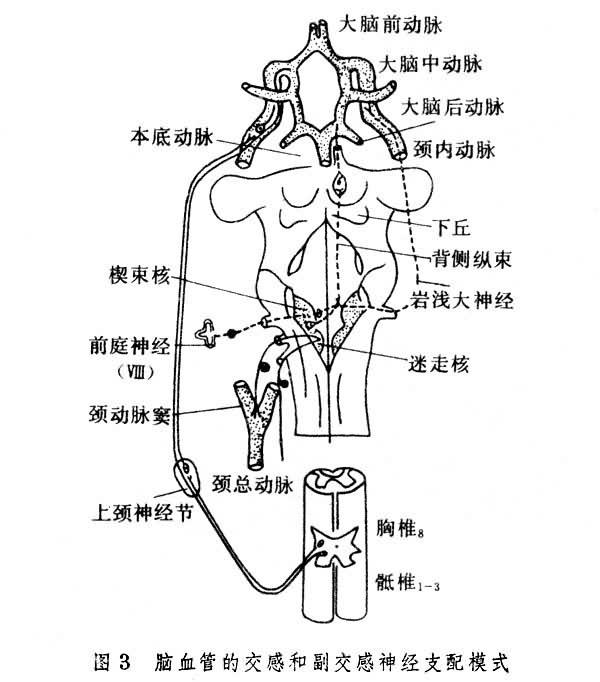 腦的白質動脈和小動脈有很多神經纖維分佈。它們來自腦幹中的去甲腎上腺素能神經元。一切中樞神經系統中灰質比白質具有更加豐富而密集的血管分佈,這與灰質較高的代謝率相適應。
腦的白質動脈和小動脈有很多神經纖維分佈。它們來自腦幹中的去甲腎上腺素能神經元。一切中樞神經系統中灰質比白質具有更加豐富而密集的血管分佈,這與灰質較高的代謝率相適應。
特征 腦循環處於堅硬顱骨腔室之內,顱腔容積是固定的,其中所含的各種組織的容積也基本固定,幾乎是不可壓縮的,例如腦組織、腦脊液和顱內血液的容積總和是接近恒定的。可是,由於圍繞腦和脊髓的神經根的硬腦膜的袋狀突出──根袖可以擴張,從而也能增加一些腦容積。
由於顱腔容積基本恒定,腦中動靜脈血管經常處於受壓狀態,因此腦血管的容積沒有搏動性變化。腦動脈沒有脈搏,血液在腦血管平穩均勻地流動著。椎動脈在進入顱腔之前發生多次的彎曲,以及腦動脈管壁與其他動脈管壁相比具有更發達的彈性纖維,也有助於腦動脈搏動的消除。其中顱腔壓迫限制作用可能更為重要,因為當顱腔密閉狀態受到破壞,如在開顱手術之後就會出現明顯的腦動脈脈搏。
腦的全部毛細血管網幾乎經常開放著,雖然有時也可以看到少許不開放的毛細血管,但沒有其他器官微循環所常見的毛細血管和前毛細血管的交替性收縮和舒張。腦毛細血管網對各種刺激的反應與內臟器官不同;在窒息狀態下,由於缺氧,全部內臟毛細血管都起收縮反應,而腦毛細血管卻起舒張反應。
正常的腦血流量 通過人工安裝的透明顱窗觀察,用流量計測大腦血流以及使用各種同位素(85氪、133氙、42鉀、137銫)作為指示劑,定時測量等方法研究全腦或局部血流,發現腦血流量之大是驚人的。如人腦雖比許多器官輕得多,但腦血流量卻占心輸出量的13~15%,腦的耗氧量也突出地高,約為3~5毫升(100克腦重每分鐘),約占全身耗氧量的18~20%。
從器官血流量和耗氧量來看,腎、肝、心肌和腦的數值都較突出,說明這些器官都很活躍。骨骼肌的血流量和耗氧量也很大,但按單位重量計算,比腎、肝、腦和心肌少得多。
血流在大腦各部的分佈也有差異。在實驗動物中,測定惰性放射性氣體在腦組織冰凍切片中的分佈,並將測定數值與血中此氣含量的比較,發現大腦和小腦皮層的血流量較大,但腦各部血流量不等;從分區來看,四疊體的下丘血流量最大(見表),灰質血流量比白質的約大6倍。
未經麻醉的貓腦各代表區的血流量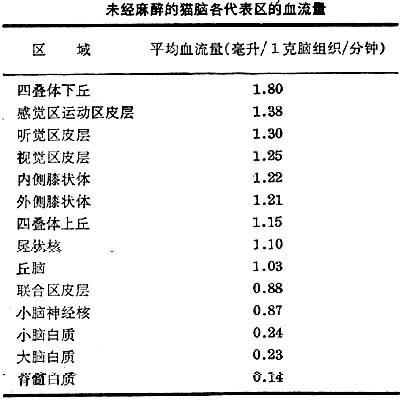
調節 腦循環受多種因素的調節,經過調節機制即使內外環境發生各種變化,大腦血流仍能保持穩定,這對腦正常機能的活動有重要意義。例如全部腦血流不因緊張的精神活動而增加,也不因精神活動的松弛乃至睡眠而減少(睡眠時甚至有些增加)。體液因素特別是腦血流中二氧化碳pH值、K+、Ca2+等對腦血管運動的調節作用比較明顯,而神經調節作用較弱,居於次要地位。腦循環因在顱腔之內,顱內壓的變化必然會有影響。此外在腦水平的平均動、靜脈壓以及血液的粘稠度都對腦循環有一定影響(圖4)。
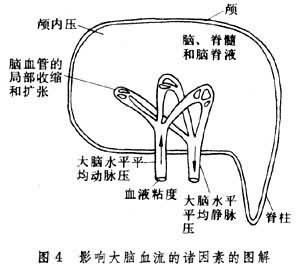
顱內壓的作用 顱內壓上升時腦血管就受到壓擠。靜脈壓的升高使靜脈血回流阻力增加會導致顱內壓上升。當顱內壓上升到450毫米水柱(33毫米汞柱)時,即使持續時間不長,腦血流也會顯著減少。這會使腦供血不足,引起血管運動中樞的興奮性提高導致全身血壓的上升。這種顱內壓升高引起的全身血壓的升高叫做庫興氏反射,有助於大腦正常血流量的保持。在一定范圍內全身血壓的升高與顱內壓的升高成正比,不過當顱內壓超過全身血壓時就會發生腦血流停止的險象。
神經調節 大腦基底部主要動脈和軟腦膜小動脈都有豐富的腎上腺素能神經纖維分佈,它們的軸突末梢有兩種囊泡;一種有顆粒的起源於交感神經,與血管收縮有關,一種無顆粒的起源於副交感神經,與血管舒張有關。腦血管一般接受腎上腺素能和膽堿能兩種神經纖維的支配,兩種神經纖維相距約250埃,這為相互作用提供瞭結構基礎。
體液調節 腦中的小動脈和身體其他器官的小動脈一樣,都直接接受局部組織氧和二氧化碳含量的影響。腦動脈二氧化碳分壓的升高具有特別顯著的血管舒張作用,使腦血流量大增。二氧化碳分壓的下降則有血管收縮作用,使腦血流量減少。通氣過度降低動脈血二氧化碳分壓,可能引起大腦癥狀。氫離子也有血管舒張作用,pH值下降引起腦血管的舒張和腦血流量的增加,pH值上升導致腦血管的收縮和腦血流的減少。事實上,二氧化碳分壓的變化引起腦中氫離子濃度的相應變化,二氧化碳的血管舒張作用是通過pH值的改變來實現的。因為大腦血流量的改變比腦組織二氧化碳分壓的變化更快。與二氧化碳分壓的緩慢改變相比,腦脊液pH值的改變要快得多。由於腦脊液的緩沖作用比血漿小得多,因此細胞外液pH值的變化比起二氧化碳分壓的變化對腦血管可能是更適宜的刺激。腦細胞外液的氫離子濃度是改變腦血管流量的重要因素但不是唯一的因素,其他如鉀離子和鈣離子也有顯著作用。鉀離子在軟腦膜小動脈周圍濃度增至10~20摩爾/升,就導致血管舒張,腦脊液鉀離子減少引起腦血管收縮。不論血管外pH值或K+濃度如何,腦血管內徑隨鈣離子的增加而減少,在無鈣離子情況下,無論pH值高低,鉀離子都會直接引起血管舒張。在腦血管調節過程中以上3種離子具有相互影響的關系。
陰離子對腦循環的調節資料很少。用缺乏氯離子的灌流液灌註腦血管周圍間隙,無論pH值大小都會引起腦血管的收縮反應。
腺苷是腦和腦脊液的正常成分,能使腦血管舒張。其作用機制是減少乃至阻斷腦血管平滑肌吸收鈣離子。腺苷直接作用於軟腦膜上的小動脈,或是使腦組織腺苷濃度增加,都能引起腦血管的舒張和腦血流的增加。腺苷、氧供應和pH值三者協同作用能顯著地增加腦血流。
氧的作用與二氧化碳的作用相反:高氧癥引起腦血管的明顯收縮,低氧癥引起腦血管明顯舒張。
腦中二氧化碳、氧和pH值對腦血流量的作用可總結為:二氧化碳分壓的增加引起腦血流量的明顯增加;氧分壓的增加作用相反。但是腦脊液和組織中細胞外液的pH值變化對腦血流的調節作用則是主要的。
兒茶酚胺對腦血流的影響很小,在整體情況下,動脈註射去甲腎上腺素後,腦血流幾乎沒有變化。不過兒茶酚胺對離體的腦血管則有明顯的收縮作用;如去甲腎上腺素、腎上腺素、異丙基腎上腺素都有這種作用。其他非兒茶酚胺的藥物如組織胺和5-羥色胺也有這種作用,不過腦血管對這些藥物的敏感性較低,反應較小。
代謝和功能調節 從1973年以來腦循環的研究趨勢是將腦血流、腦代謝和腦功能結合起來進行研究,這與腦局部血流測定方法學上的改進具有密切關系。
正常情況下,腦的局部血流是由神經和局部代謝調節的。外界環境多種刺激、身體活動和意識活動增加都會使大腦相應代表區局部血流量增加,如手部強烈運動時大腦皮層手部代表區的局部血流量明顯增多,隨著活動加強,血流增加區域沿著中央溝擴展。感覺沖動增多,包括中等程度的痛覺會使中央前回比後回有較多的血流量增加。隨意運動時,運動後區比前區血流增加得更多。
電刺激大腦不同部位也可明顯影響腦的局部血流。例如電刺激中腦被蓋的網狀結構和海馬,可使中隔部血流量增加。電刺激一側海馬使其先有癲癇樣放電,繼而出現中隔血流的成倍增加。
參考書目
W.F.Ganong,Review of Medical Physiology,10thed.,Lange Medical Publications,Los Altos,1981.
V.B.Mountcastle,Medical Physiology,14th ed.,Mosby Co.,St.Louis,1980.