━━━━━━━━━━━━━━━━━━━━━━━━
疏鬆結締組織
纖維
基質
細胞
脂肪組織
網狀組織
緻密結締組織
彈性組織
軟骨
骨組織
結締組織的發生
膜內成骨
軟骨內成骨
結締組織的比較組織學
━━━━━━━━━━━━━━━━━━━━━━━━
多細胞動物體內起連接和支持作用的基本組織,其形態特征是:細胞類型較多,且細胞外存在有大量由纖維和基質組成的細胞間質。結締組織可分為幾種類型。機體器官內外廣泛存在的是疏松結締組織,其中纖維較少且排列松散,基質中包容著大部分細胞外液,代謝物質可在其間流動。與疏松結締組織連接的組織在一定范圍內可相互移動。其中的細胞還有免疫防禦和組織修復等功能。有的細胞貯存營養;脂肪組織便是特種的疏松結締組織。造血組織(骨髓、淋巴)的支架是由網狀纖維組成的網絡結構;這種網狀組織也屬於疏松結締組織。致密結締組織中,纖維較多,且緊密交織在一起,可承受較大的負荷。纖維或交織成網,如被覆體表的真皮或包繞器官的被膜;或並聯成束,如連接骨與肌肉的肌腱或連結骨與骨之間的韌帶。軟骨和硬骨則為致密結締組織中最堅實的。其中纖維排列最為致密,軟骨中的基質形成堅實的凝膠,硬骨中更沉積瞭大量鈣鹽。因此它們成為身體的主要支架、運動的杠桿以及保護中樞神經系統和內臟的外殼。
在種系發生中,結締組織的出現遲於上皮組織,而且大部是在脊椎動物中才出現的。在個體發生中,結締組織都來自發源於中胚層的間充質。工業上應用的皮膠便是由動物結締組織中提取的膠原蛋白。結締組織在醫學上也很受重視,隨著機體的老化,膠原纖維間交叉聯結增多,基質含水量減少,從而可能影響代謝物質的運轉。疏松結締組織散佈機體各處,大量的小血管和毛細血管走行其間,結締組織還富含免疫和炎性細胞,因而有所謂結締組織病(舊稱膠原病)發生於此。不過,其中大部分實為過敏性脈管炎,並非源起於結締組織本身。
疏松結締組織
疏松結締組織散在機體各處的器官之間、組織之間甚至細胞之間。纖維較少,基質較多,散佈於基質中的細胞類型最多,因此除瞭連接、支持等作用外,它還具有免疫防禦、組織修復和貯藏營養等功能(圖1)。
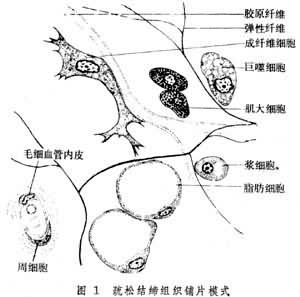
纖維 有3種,以膠原纖維最多,彈性纖維次之,網狀纖維最少。
膠原纖維 主要由Ⅰ型膠原蛋白組成(見膠原和明膠)。纖維直徑1~20微米,有分支,呈波狀走行,互相交織成網。電子顯微鏡下,膠原纖維由更細的膠原原纖維集合而成,膠原原纖維的直徑約20~90納米,有明暗交替的橫紋。膠原纖維的韌性大,抗拉力強。
彈性纖維 主要由彈性蛋白組成,一般細於膠原纖維,無原纖維,有分支,常與膠原纖維交織一起(圖1)。電鏡下,彈性纖維的主體呈均質狀;為彈性蛋白部分,其周圍有直徑10納米,由糖蛋白組成的細管狀微纖維。彈性纖維的彈性大。
網狀纖維 由Ⅲ型膠原蛋白組成,纖維較細,直徑0.5~2微米,分支多,連結成網。電鏡下,網狀纖維由更細的原纖維集合而成,這種原纖維的直徑約45納米,呈與膠原原纖維同樣的周期性橫紋。
基質 是充填在纖維和細胞周圍的間隙內的均質膠狀物。基質除含有水、無機鹽等外,其主要成分是葡氨聚糖和一些糖蛋白。近百個含硫酸的葡氨聚糖線性分子結合在一核心蛋白長鏈上,構成試管刷樣的蛋白聚糖結構。蛋白聚糖能結合大量水分子,使基質呈膠體,有支持其間細胞的作用。基質中含有組織液,組織液是血液與組織細胞間交換代謝物質的中間環節,而各器官內的毛細血管一般均勻分佈在疏松結締組織中,故結締組織基質的通透性對細胞代謝有重要關系。
細胞 以成纖維細胞為主;巨噬細胞也較多,漿細胞及肥大細胞則很少,這後3種細胞均屬於免疫防禦細胞,它們都來自造血組織。
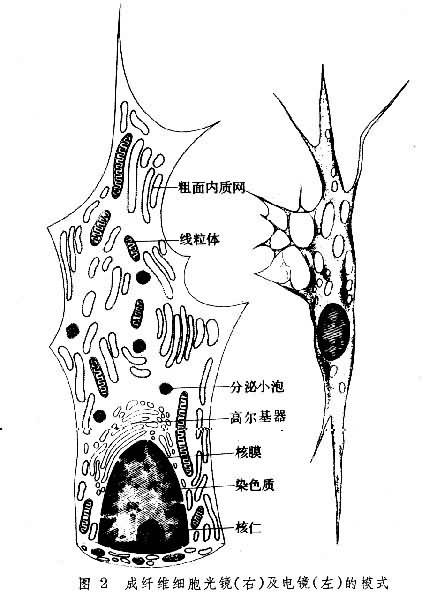
成纖維細胞 常附於膠原纖維一側(圖1)。其形態結構隨其功能狀態而異。功能活躍的成纖維細胞有較多突起;胞核較大,核仁明顯;胞質豐富,呈嗜堿性;電鏡下,呈顯蛋白分泌細胞的特征(圖2)。靜止的成纖維細胞又稱纖維細胞,細胞較小,呈梭形,胞核小而染色較深,胞質嗜酸性,其中的細胞器較少。成纖維細胞是形成細胞間質的主要細胞。它合成並分泌成各種纖維的蛋白質,這些蛋白質分泌到細胞外後分別聚合成不同的纖維;它也合成並分泌葡氨聚糖、蛋白聚糖、糖蛋白等基質成分。在組織生長和創傷修復時,成纖維細胞功能活躍,積極進行細胞間質的形成;平時,則參與細胞間質的代謝更新。
巨噬細胞 由遊出血管的單核細胞衍變而成,其體積較大,形狀不規則,常伸出短而鈍的突起(圖1)。電鏡下,胞質內有很多溶酶體。巨噬細胞能作變形運動,能吞噬衰老、死亡的細胞和侵入體內的異物、細菌,形成吞噬體。吞噬體及吞飲小泡與初級溶酶體融合形成次級溶酶體,其內容物被溶酶體酶分解消化。故巨噬細胞有重要的防禦保護作用。巨噬細胞還是免疫細胞的成員之一,在免疫反應中起重要作用。
漿細胞 由 B淋巴細胞在抗原刺激下衍變而成,在一般疏松結締組織內罕見,多見於經常受抗原刺激的消化道、呼吸道的固有膜結締組織中(見淋巴細胞)。
肥大細胞 多沿小血管分佈,體積較大,呈卵圓形或圓形,胞質充滿有異染性的嗜堿性顆粒(圖1)。顆粒含肝素、組胺、嗜酸粒細胞趨化因子等物質,有些動物如大鼠的顆粒還含5-羥色胺。但對肥大細胞的生理意義仍不十分明瞭。肥大細胞表面有免疫球蛋白E(IgE)受體。抗原初次進入機體引起漿細胞產生的IgE,大部分結合在肥大細胞(及嗜堿粒細胞)表面。當同樣抗原再次進入機體時,就與肥大細胞表面的IgE結合,導致細胞釋放顆粒。顆粒內容物釋放後引起炎癥反應。一般來講這種反應可以清除抗原及消除其危害,於機體有益。但對某些個體,再接觸同樣抗原即使肥大細胞迅速大量釋放顆粒,引起速發型過敏反應(如支氣管哮喘、青黴素過敏性休克)。
此外,疏松結締組織還有散在或成群的脂肪細胞(圖1)、從血管遊出的少數白細胞及未分化的間充質細胞。這後一種細胞多分佈在毛細血管附近,形態似成纖維細胞。它保持胚胎間充質細胞的分化潛能,在結締組織再生時能分化為成纖維細胞,血管再生時能分化為血管壁的平滑肌細胞。
脂肪組織 分佈在皮下、腹膜後、網膜和腸系膜等部位,由大量脂肪細胞群集而成,被一般疏松結締組織分隔成許多脂肪小葉。脂肪細胞很大,直徑可達120微米。呈球形或多邊形,脂肪聚成大滴,占據細胞絕大部分,胞質、胞核被其擠到細胞邊緣。在一般染色切片上,由於脂肪被溶去,脂肪細胞呈戒指狀。脂肪組織的主要功能包括:貯存能量;隔熱保溫;固定器官的位置;以及在手掌、足底形成脂肪墊,起減震器的作用。
上述脂肪組織在體內分佈廣,呈白色或黃色,又稱白色脂肪組織。另有棕色脂肪組織,主要存在於胚胎和幼年哺乳動物體內局部部位,成年動物中隻有冬眠種類的棕色脂肪特別發達,這種脂肪組織的脂肪細胞較小,胞核位居中央,胞核周圍有許多大小不一的脂滴。這種組織中貯存的脂肪,氧化時隻能化為熱能,在冬眠動物復溫時起重要作用。
網狀組織 由網狀細胞及其產生的網狀纖維組成。星狀多突的網狀細胞互相借助於突起相連,依附在網狀纖維上,並由其胞突包裹網狀纖維,構成網狀支架(圖3),成為骨髓組織和淋巴組織的支持組織。

致密結締組織
致密結締組織的組成成分與疏松結締組織的基本相同,特點是基質較少而纖維排列緊密,故支持連接功能強。依纖維的排列方式,一般致密結締組織常分為規則和不規則的兩型。不規則的致密結締組織,構成真皮、肝、脾等器官的被膜,軟、硬骨膜及硬腦脊膜等。纖維以膠原纖維為主,也有彈性纖維。纖維的走向不一,互相緊密交織在一起。在纖維之間有細胞,以成纖維細胞為主。此種組織的韌性很大,主要起保護作用。骨外膜和軟骨膜的內層有些細胞尚未完全分化,它們參與骨和軟骨的形成。
規則的致密結締組織是構成肌腱、韌帶的主要組織,以肌腱最為典型。肌腱主要由粗大的膠原纖維束密集平行排列而成,纖維束間有少量基質。成纖維細胞在纖維束間成行排列,細胞伸出多個薄翼狀突起包圍纖維束。腱的膠原纖維一端與肌肉的肌纖維相連,另一端埋在骨內。腱的承受很大的牽引力,並能將肌內收縮的牽引力傳遞到骨,以牽動骨關節運動。
彈性組織 是由彈性纖維組成的規則致密結締組織,見於心臟、大動脈、呼吸道以及項韌帶等少數韌帶和馬等大型傢畜的腹壁筋膜。
軟骨 由軟骨細胞和細胞間質組成。軟骨的細胞間質,又稱軟骨間質,含有纖維和凝膠狀基質,基質含水量大,堅韌且具彈性,故軟骨有吸震、潤滑等功能。軟骨間質內有許多分散的腔隙,稱為軟骨陷窩,軟骨細胞就位於陷窩內。軟骨細胞有形成纖維和基質功能。細胞周圍的基質含葡氨聚糖較多,呈強嗜堿性,稱軟骨囊。軟骨內一般沒有血管,由四周軟骨膜內的毛細血管,通過軟骨基質中的組織液為軟骨細胞提供營養。軟骨膜由致密結締組織構成,被覆在軟骨表面(除關節軟骨的關節面和纖維軟骨外)。其內層有些細胞尚未完全分化,它們能分裂增生,形成軟骨。軟骨分為以下3種:
透明軟骨 分佈最廣,構成呼吸道的軟骨(個別除外)、肋軟骨和關節軟骨。軟骨中膠原纖維主要由Ⅱ型膠原蛋白構成。基質約含60%的水,呈凝膠狀。軟骨基質中的蛋白聚糖常又組成更大的復合物。多個蛋白聚糖的核心蛋白各通過一個連接蛋白結合到一個透明質酸分子上,透明質酸也是葡氨聚糖的一種,但它很長,旦不結合硫酸根。一個透明質酸分子可以結合幾十個蛋白聚糖組成一個更大的試管刷樣結構。具硫酸葡氨聚糖借硫酸根與膠原原纖維靜電結合,並結合大量水。以關節面軟骨為例,它的潤滑及吸震性能,正是由於這種含水結構造成的。近軟骨表面的軟骨細胞幼稚,體積小,呈扁橢圓形。向軟骨深部,軟骨細胞變大變圓,常2、4或8個細胞成群分佈,由於每群細胞是由一個幼稚軟骨細胞分裂而成,故稱同源細胞群。
彈性軟骨 分佈於耳廓、外耳道、咽鼓管及會厭,並構成少數喉軟骨。此種軟骨的構造與透明軟骨基本相同,特點是在軟骨間質內有大量彈性纖維,故此種軟骨既堅韌又富於彈性。
纖維軟骨 分佈在椎間盤、恥骨連合、以及肌腱的韌帶附著於骨的部位等處,具有加強連接的作用。其結構特點是:膠原纖維束平行排列,在纖維束之間有排成單行的軟骨細胞,基質少。纖維軟骨沒有軟骨膜,由致密結締組織的成纖維細胞轉變為軟骨細胞。
骨組織 最堅硬的一種結締組織,是構成全身硬骨(bones)的基本成分,具有重要的支持、運動和保護作用。體內的鈣約99%以磷酸鹽形式貯存在骨組織內,故骨組織對體內鈣磷代謝起重要作用。骨組織由骨細胞和細胞間質組成。骨的細胞間質,又稱骨間質,包含有機物和無機物。有機物中,膠原纖維(主要Ⅰ型膠原蛋白組成)占95%,其餘為葡氨聚糖及蛋白質。無機物占骨的幹重的50%,以鈣、磷為最多,它們形成羥基磷灰石結晶。電鏡下結晶呈細針狀(40×25×3納米),分佈在膠原原纖維旁。除個別骨外,成年哺乳動物骨組織的膠原纖維均平行排列成板層狀,由有機基質粘合一起,並有骨鹽沉著。這種薄板狀結構稱為骨板(圖4)。相鄰兩層骨板的纖維走向互成一定角度,這有利於增強骨的堅固性。在骨板之間和骨板內有散在的扁橢圓形小腔,叫骨陷窩。骨陷窩發出許多輻射狀的小管,叫骨小管,與鄰近的骨陷窩相通。骨細胞位於骨陷窩內,它有許多長突起伸到骨小管中,與相鄰骨細胞的突起接觸,形成縫隙連接。骨間質有大量骨鹽沉著,組織液不能從其中擴散,但骨陷窩-骨小管系統中存在骨液,可從鄰近的血管獲得營養傳遞給骨細胞,並參與血鈣濃度的調節。骨細胞之間還可借縫隙連接交換物質。根據形態結構的不同,骨組織分為松質骨和密質骨。松質骨,分佈在長骨(如肱骨、股骨)兩端即骨骺、和短骨(如椎骨)及扁骨(如肩胛骨)內部。由數層至十餘層骨板構成針狀、片狀骨小梁,骨小梁相連成海綿狀,在骨小梁間的間隙內充滿紅骨髓,有豐富的血管。密質骨,見於長骨的骨幹,長骨兩端骨骺表面、短骨和扁骨表面,骨板密集,有4種排列方式,如長骨幹(圖4);①外環骨板,位於骨幹外表面,是數至十數層與骨幹表面平行的骨板;②內環骨板,位於骨幹骨密質內表面,約有數層骨板;③哈弗斯氏骨板,位於內、外環骨板之間,由數至十數層圓筒狀骨板同心排列而成,順骨幹長軸平行配佈。在哈弗斯氏骨板中央有一條縱行管道,稱哈弗斯氏管,內有血管、神經等結構。哈弗斯氏骨板和哈弗斯氏管合稱哈弗斯氏系統,也稱骨單位;④間骨板,填充在哈弗斯氏系統之間,是被破壞的哈弗斯氏系統所殘留的一些骨板。密質骨的骨板排列方式,保證瞭骨的強度,也保證瞭深在的骨細胞,如哈弗斯氏骨板中的骨細胞獲得營養。
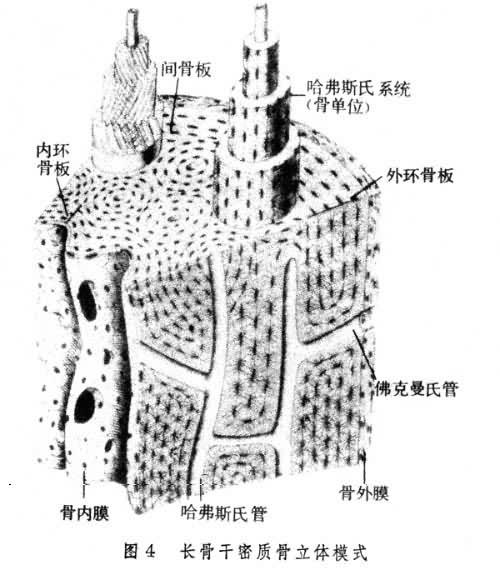
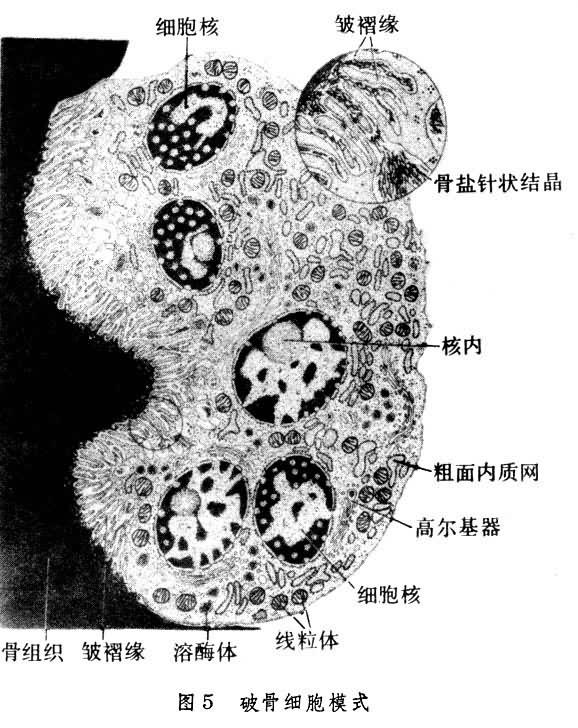
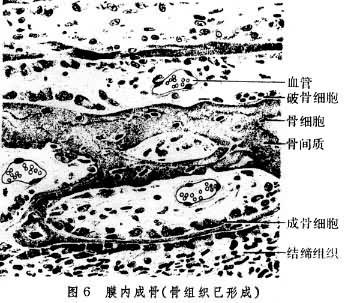
在骨組織表面被覆著骨膜,分為骨外膜和骨內膜。骨外膜,覆於各骨的外表面(但關節面無骨膜),由較厚的致密結締組織構成。骨內膜為薄層結締組織膜,覆於松質骨的骨小梁表面、密質骨的骨髓腔面及哈弗斯氏管表面。骨膜的血管營養骨組織。骨膜還對骨的生長、改建以及骨折的愈合起重要作用。在骨外膜深層及骨內膜有分化較低的骨原細胞,它的形狀與成纖維細胞相似,當骨組織形成時,它變得活躍,進行分裂增生,並分化為成骨細胞進行造骨。成骨細胞呈立方或矮柱狀,有突起,成骨活躍時常排成單層貼附於骨組織表面(圖6)。成骨細胞合成並分泌有機骨間質,並參與骨鹽沉著,形成新骨組織;成骨細胞被埋入骨間質後,就變為骨細胞。此外,在骨組織進行吸收部位,常見破骨細胞附於骨組織邊緣。它是由單核細胞融合而成的多核大細胞(圖5)。破骨細胞參與骨間質的溶解吸收,故多位於骨質吸收後所形成的淺凹處。
結締組織的發生
結締組織來源於胚胎早期的間充質。間充質遍存於3個胚層之間,正在發生的器官內外。間充質來自中胚層,由間充質細胞和柔軟膠狀細胞間質組成。在結締組織發生時,間充質細胞分化為成纖維細胞、成脂肪細胞、網狀細胞、成軟骨細胞,成骨細胞,這些細胞產生相應的細胞間質,分別形成各種纖維性結締組織、脂肪組織、軟骨和骨組織。卵黃囊血島的間充質細胞分化為造血幹細胞,經血管遷至造血器官,產生各種血細胞。骨的發生和生長是個長而復雜的過程。骨的發生有膜內成骨和軟骨內成骨兩種方式。
膜內成骨 也稱膜內骨化,是顱骨的一些扁骨及面骨的成骨方式,在即將成骨處的間充質先分化為富於血管的胚胎性結締組織膜,成骨過程就在其中進行。其中一些間充質細胞分化為成骨細胞,細胞間以突起互相連接,成骨細胞產生有機骨間質,細胞本身被埋在間質內,變為骨細胞,這種初成的組織內尚無骨鹽沉著,稱為類骨質。隨後,大量骨鹽沉著在類骨間質中,即成為骨組織。骨組織開始發生的部位稱為骨化中心,所形成的骨組織呈針、片狀,漸相連成海綿狀。成骨過程由骨化中心向四周擴展,骨的容積逐漸增大(圖6)。其表面的間充質分化為骨膜,繼續在原始松質骨表面造骨,使骨漸大漸厚。骨經過不斷的生長與改建,終為成體的骨。魚類的真皮磷片及顱的真皮是進化中最早出現的膜內成骨。
軟骨內成骨 也稱軟骨內骨化,是除上述扁骨及面骨外身體內大部分骨的成骨方式。以人的長骨為例,在將成骨部位,間充質首先分化形成一塊透明軟骨,稱軟骨雛形,以後成骨過程就在軟骨表面及內部進行。當軟骨發育到相當程度,軟骨中部(相當未來的骨幹)的軟骨膜內成骨方式形成骨組織,環繞軟骨中部周圍,稱為骨領;此後軟骨膜改稱骨外膜,負責骨領的生長。不久,軟骨中部內的軟骨間質鈣化,軟骨細胞死亡,血管攜同間充質細胞、成骨細胞、破骨細胞等自骨外膜穿過骨領侵入破骨細胞將鈣化軟骨間質溶解,形成許多不規則的腔隙,成骨細胞在殘存的鈣化軟骨間質表面形成類骨質,再經骨鹽沉著而為骨組織。這個骨化中心稱為初級骨化中心。初級骨化中心所成的骨小梁相繼被吸收,在骨幹中央形成骨髓腔。以後,骨幹兩端的軟骨繼續生長,初級骨化中心的成骨過程也由骨幹向兩端推進,使骨不斷長長;骨外膜也繼續成骨,使骨逐漸變粗至出生前後,骨幹兩端的軟骨內先後出現次級骨化中心,形成骨骺的骨小梁和松質骨。隨著骺部表面的軟骨向四周生長,次級骨化中心成骨過程也向四周擴展,使骨骺增大。在骨骺表面的軟骨終生保留;成為關節軟骨。在骨骺和骨幹之間,也保留一片軟骨,稱為骺板。骺板的存在對長骨繼續長長具有重要作用。到17~20歲左右,骺板消失,骨骺與骨幹的骨組織連成一體,此後長骨即停止加長。骨的增粗約在25~30歲左右停止。
結締組織的比較組織學
結締組織是隨著中胚層的出現而形成的,並在動物進化中發展的。海綿動物和腔腸動物沒有中胚層,在它們的內、外胚層之間有一層由膠原樣纖維和葡氨聚糖多糖基質構成的中膠層,這是外胚層表皮的上皮肌細胞的產物。扁形動物有瞭中胚層,出現中胚層衍生的間充質充填在體壁與腸壁之間。在無脊椎動物的進化中,出現瞭血液,形成瞭由成纖維細胞、膠原纖維和基質構成的結締組織,並逐漸出現其他類型的結締組織。頭足動物如烏賊等,有一種特殊的軟骨,它的軟骨細胞有許多分支的長突起,與脊椎動物的骨細胞相似。舌貝(腕足動物)觸冠裡的軟骨樣組織,與脊椎動物的透明軟骨相似。昆蟲的結締組織較發達,出現由不同類型脂肪細胞構成的脂肪體,某些昆蟲的造血組織內有網狀細胞,甚至還有網狀纖維。棘皮動物如海星、海膽、海參等的體壁真皮內,有由結締組織細胞分泌物形成的含石灰質的骨片。結締組織在脊椎動物進化中得到更大的發展,類型很多,組成成分也愈益多樣。結締組織的多樣性和結構的復雜性,適應機體功能的多方面的需要。中胚層生成的內骨骼取代無脊椎動物由表皮衍生的外骨骼,從內部支持機體更便於脊椎動物生長和運動。圓口類和軟骨魚的內骨骼由軟骨構成,一些硬骨魚、兩棲動物、爬行動物、鳥類及哺乳動物的內骨骼主要由骨組織構成。內骨骼在脊椎動物進化過程中變得更為堅硬,支持作用更強。這種進化在骨組織類型方面也有反映。根據纖維束的粗細和排列方式,骨組織分為幾型,分別見於不同進化等級的脊椎動物,平行粗纖維束骨見於魚類和兩棲動物,編織粗纖維束骨見於兩棲動物和爬行動物,平行細纖維束骨見於鳥類長骨,而哺乳動物的骨主要為板層細纖維束骨。在一定容積下,板層骨的強度比編織骨的更大。板層骨取代編織骨,使哺乳動物在支持和運動功能上更為優越,是適應陸上生活的更高階段。